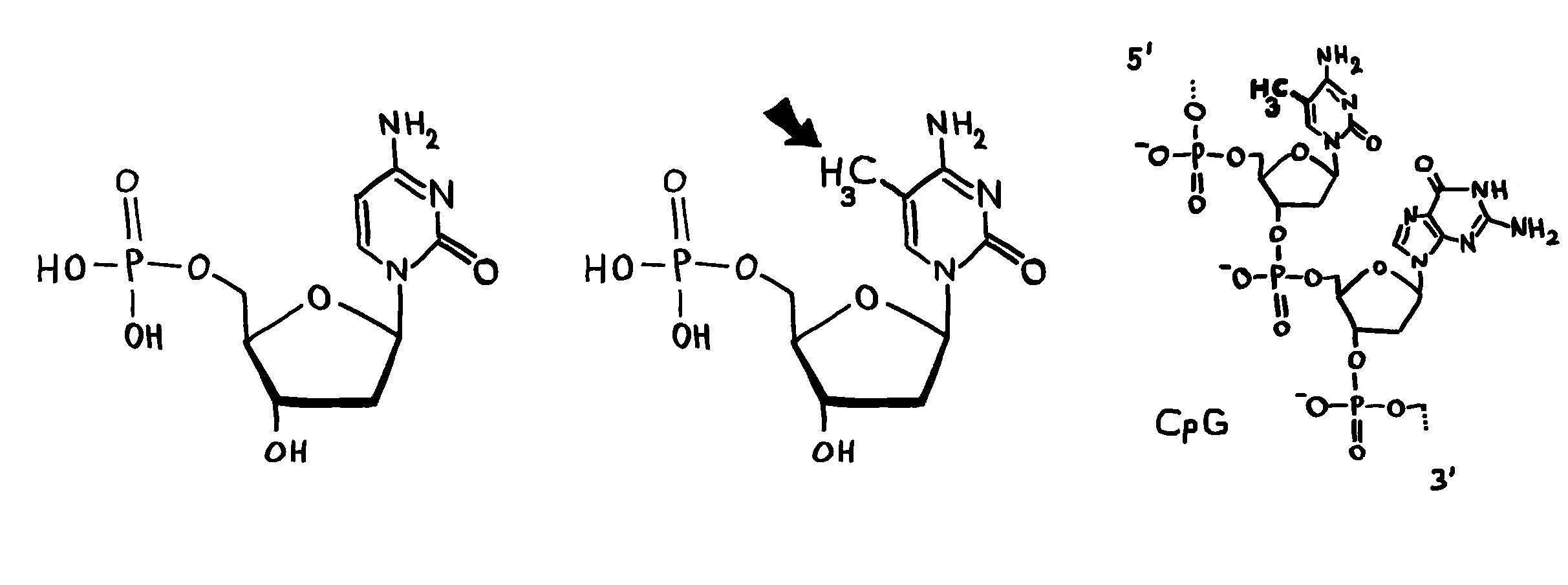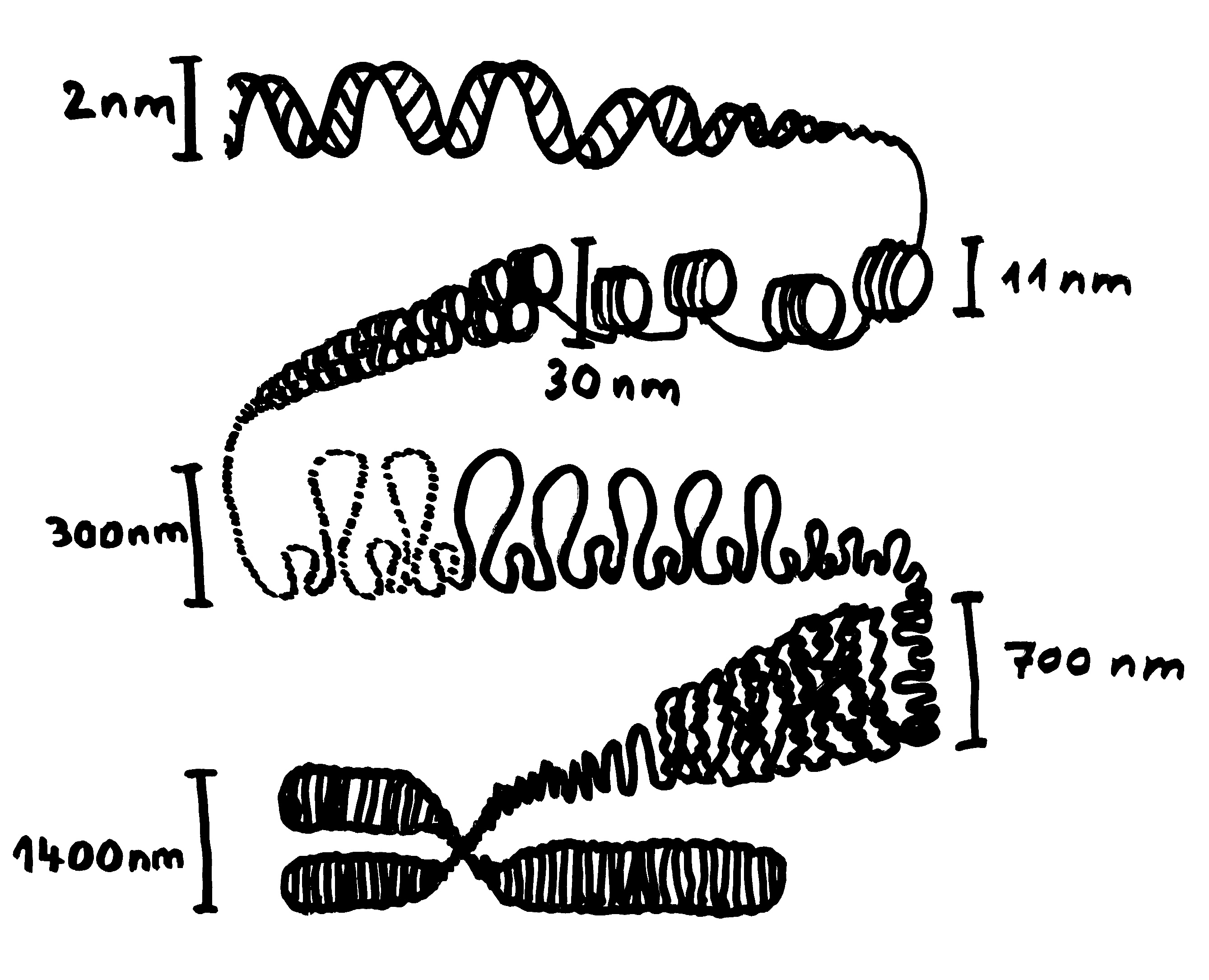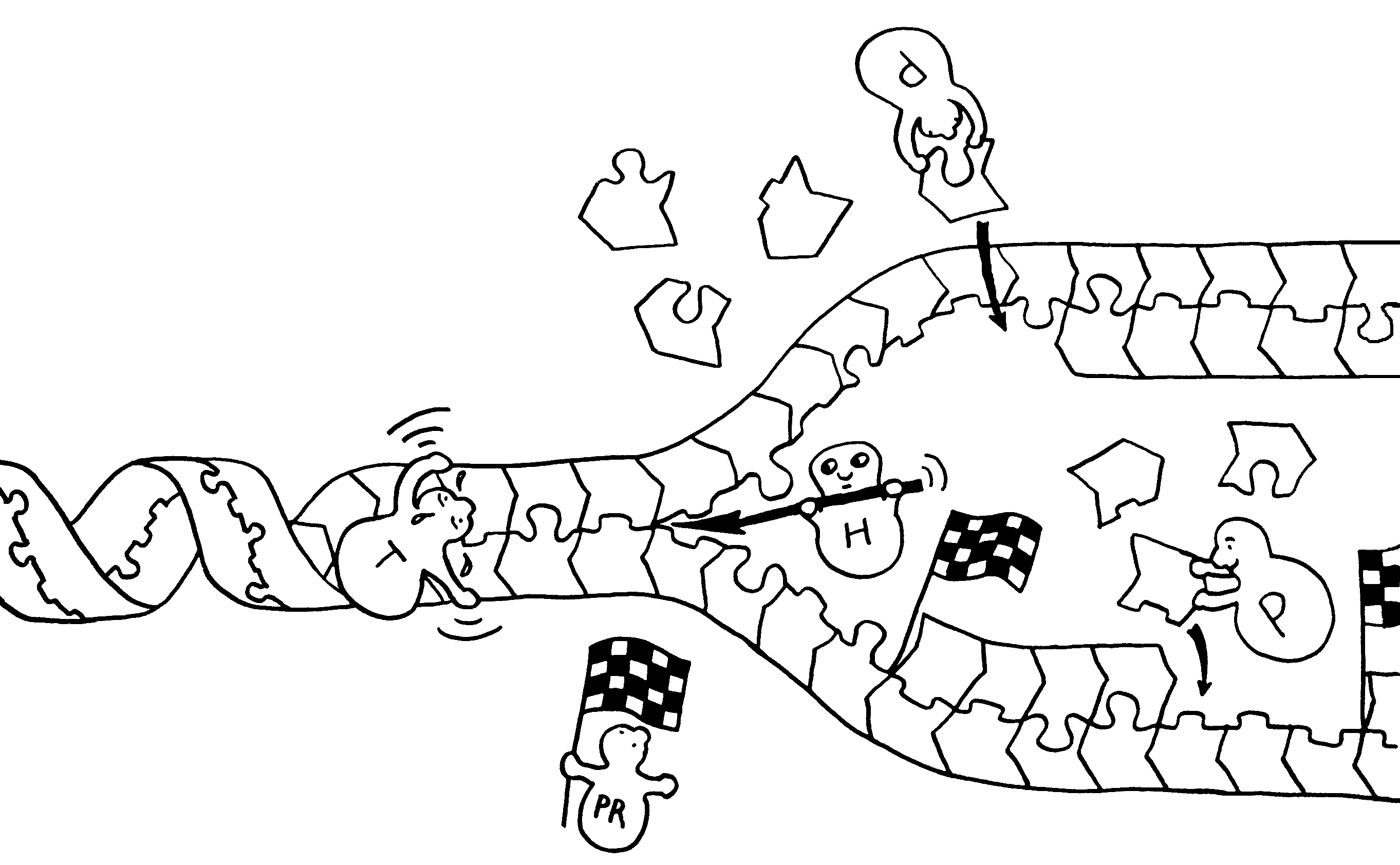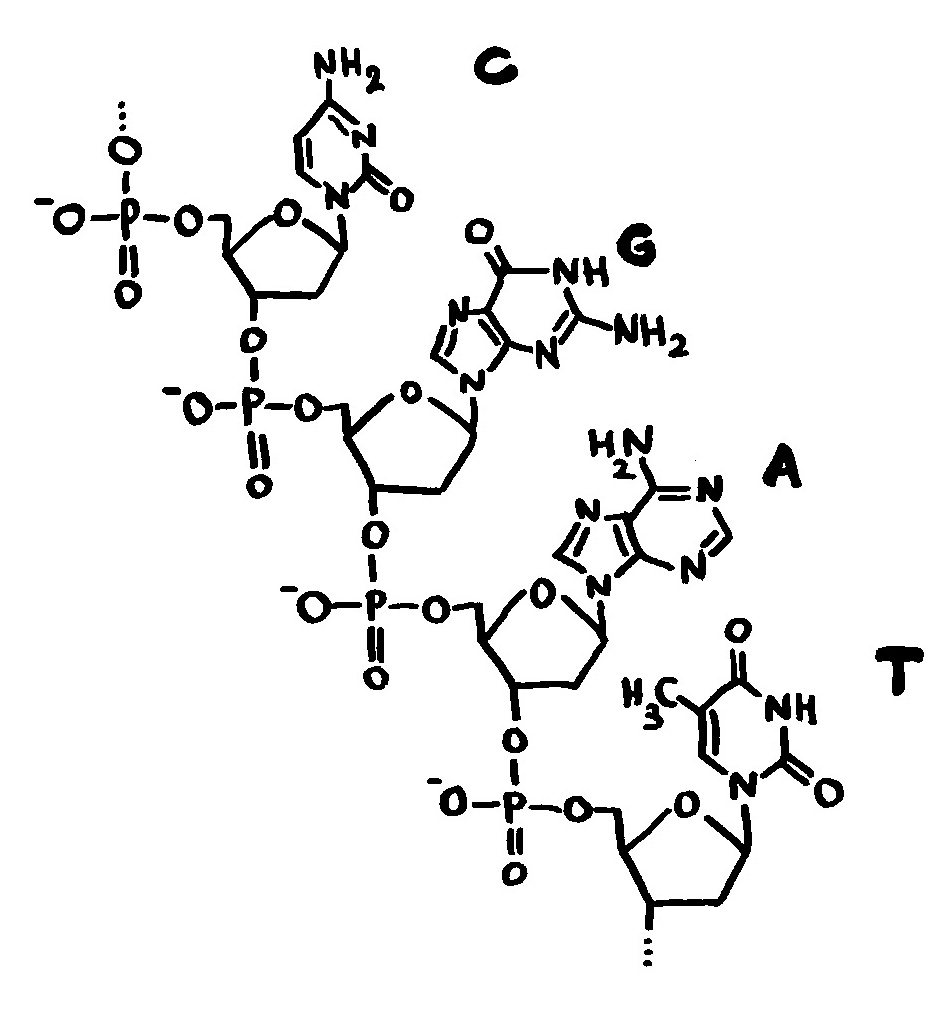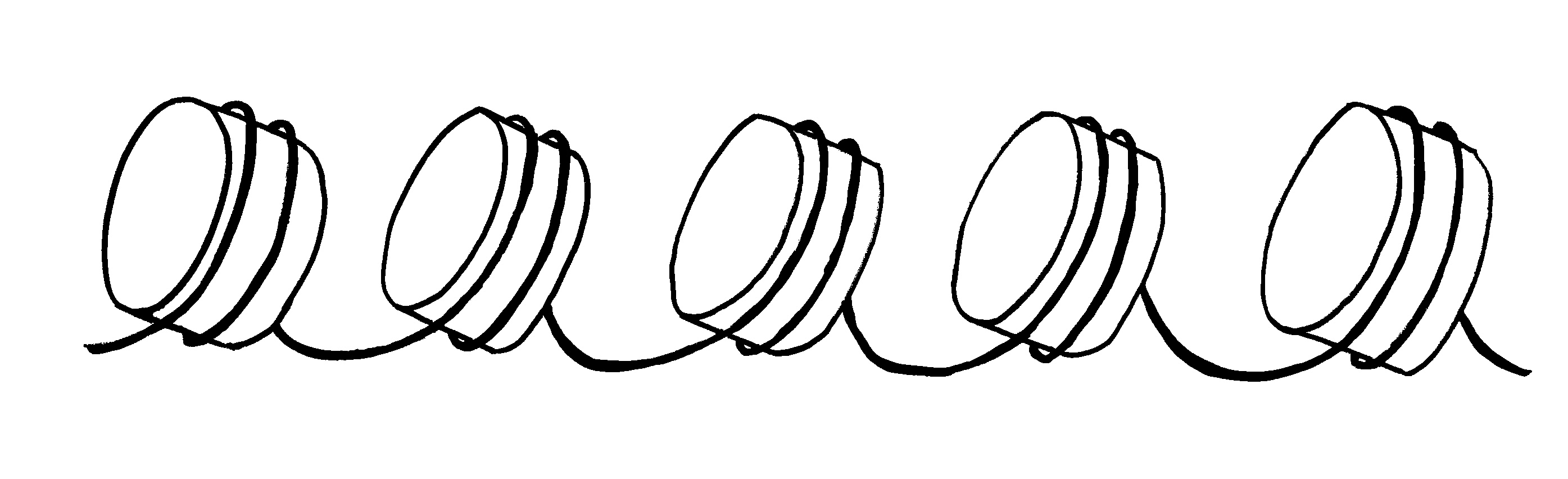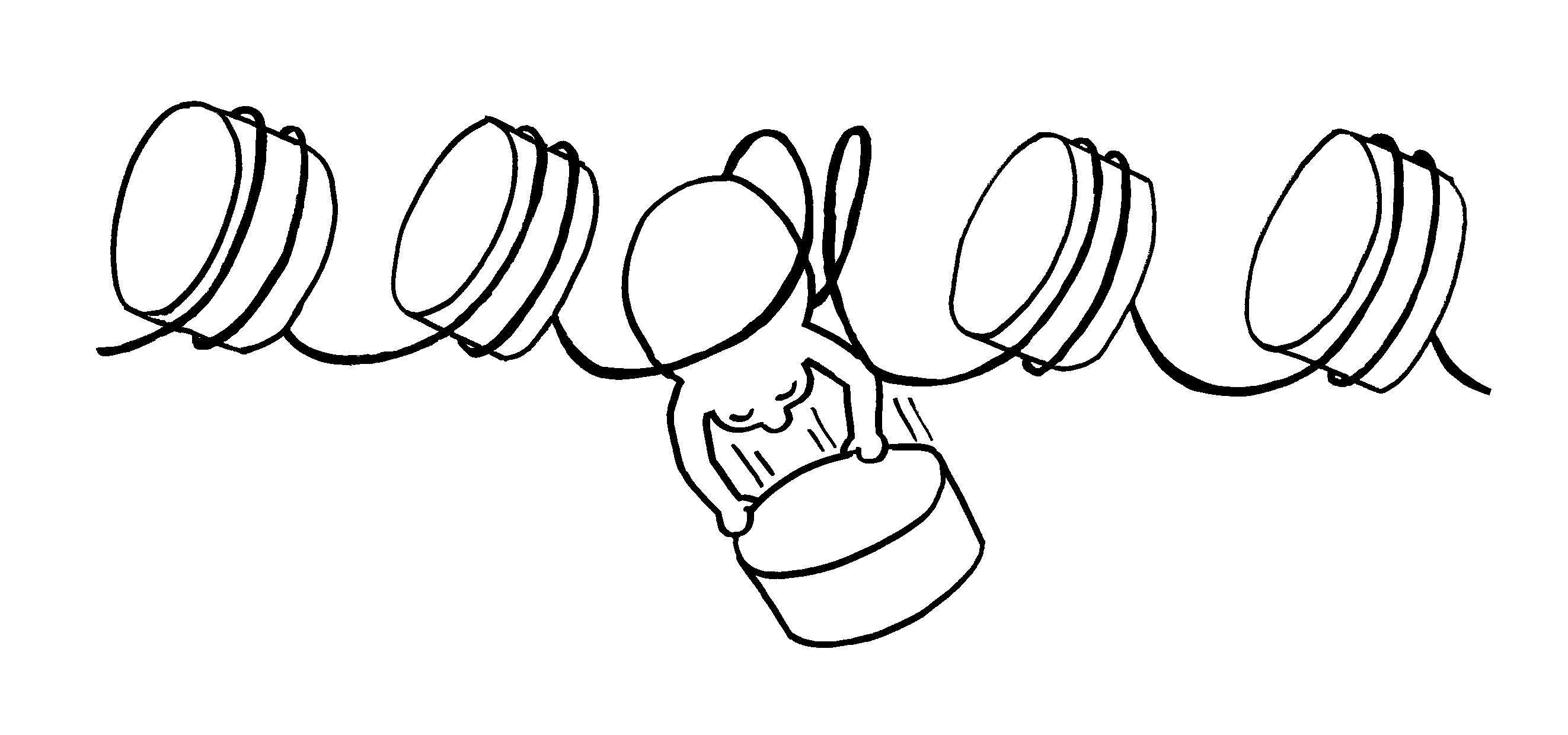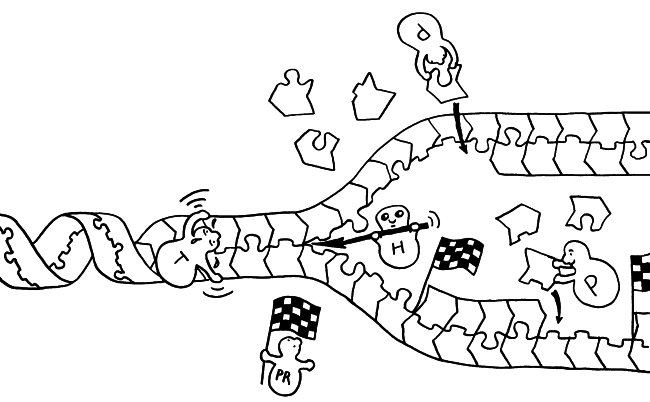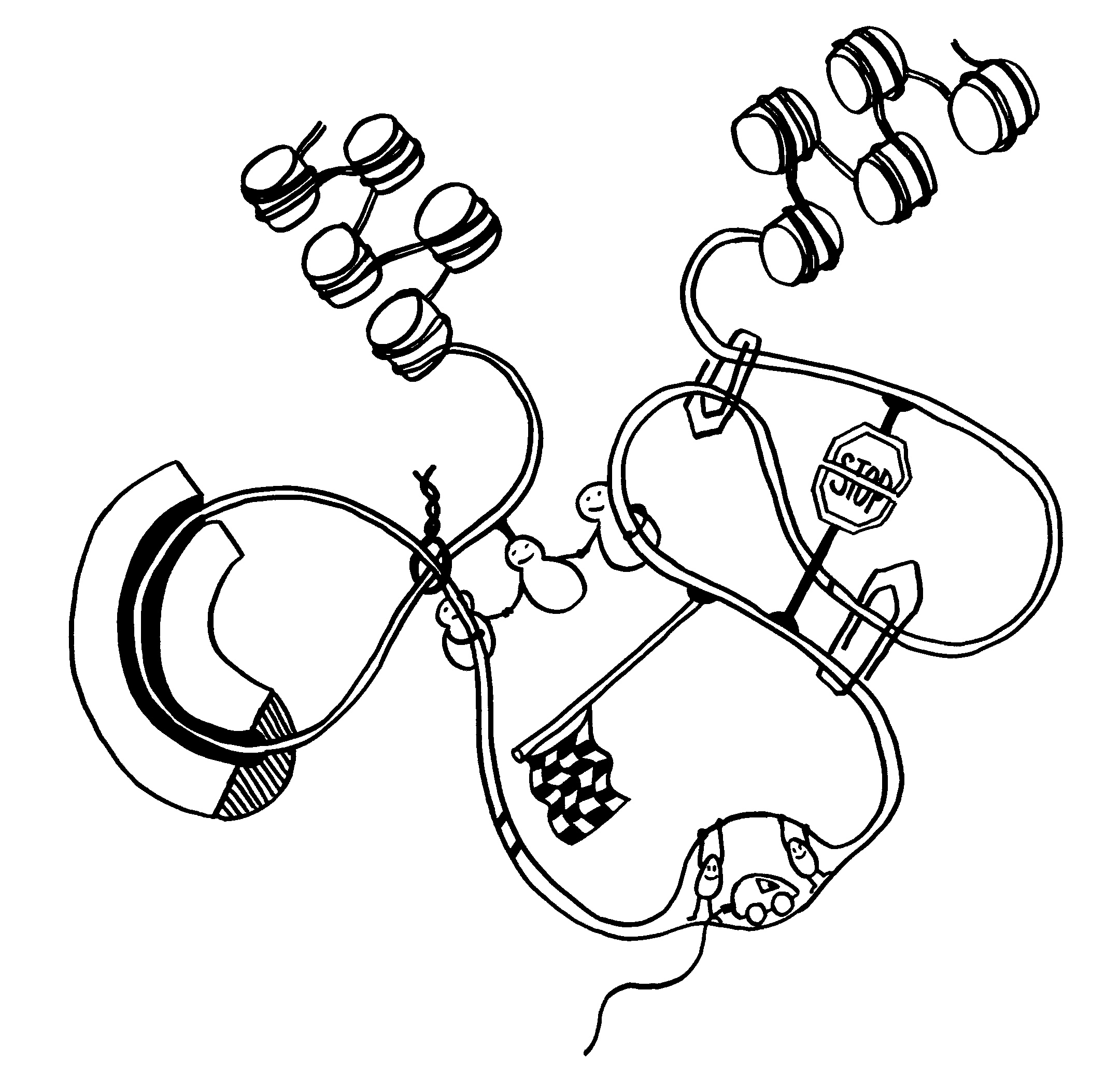
Ob ein Gen abgelesen wird und wie stark, hängt nicht nur von den DNA-Sequenzen in seiner Promotor-Region ab, an die die Transkriptionsmaschinerie andockt (Auto mit mRNA-Schwänzchen). Die DNA muss auch dekondensieren, sich also von den Nukleosomen (Zylinder im Hintergrund) lösen und zum lockeren Faden werden. Oft steuern auch sogenannte Enhancer
aus größerer Entfernung die Gen-Ablesung. Diese Fernsteuerung erfolgt über Proteine, die sich an die DNA anlagern und sie krümmen (Rinne links), in Schlingen legen (Figürchen in der Mitte) oder zusammenfalten (Büroklammern rechts). Dadurch gelangen DNA-gebundene Signale in die Nähe der zu beeinflussenden DNA-Sequenz (Startfahne und Stopzeichen).
Kleine Varianten in Steuerungssequenzen können das Risiko von Autoimmunerkrankungen
erhöhen, indem sie die Transkription eines Gens erleichtern oder erschweren. Wegen der oft beträchtlichen Entfernung zwischen Steuerungssequenz und Wirkungsort sind solche Zusammenhänge nicht leicht nachzuzweisen.
Sie dürfen diese Zeichnung gerne in Folien etc. übernehmen, sofern Sie die Quelle angeben: Dr. Andrea Kamphuis, https://autoimmunbuch.de