Unser Gehirn ist ein immunologisch privilegiertes Organ, in dem Immunreaktionen besonders strikt reguliert werden, um Kollateralschäden zu vermeiden. Dennoch enthält es Immunzellen, vor allem solche der angeborenen Abwehr – insbesondere Mikroglia.
Zu deren Aufgaben gehört das Pruning: das Wegschneiden überflüssiger Verbindungen (Synapsen) zwischen Nervenzellen, vor allem während der Kindheit und Adoleszenz. Mikroglia sind gewissermaßen die Gärtner des Gehirns, die die Sträucher regelmäßig zurückschneiden, bevor sie zu einem undurchdringlichen, dysfunktionalen Gestrüpp zusammenwuchern. Krankhaft überaktive Mikroglia übertreiben das Stutzen; sie zerstören auch Verbindungen zwischen Neuronen, die für die Gehirnfunktion notwendig sind. Andererseits sind auch erschöpfte, nicht hinreichend aktive Mikroglia schädlich, denn sie kommen mit dem Aufräumen, dem Entsorgen von Krankheitskeimen oder toten oder erkrankten Nervenbestandteilen nicht mehr hinterher. Abnorme Mikroglia werden unter anderem mit Multipler Sklerose, aber auch mit Alzheimer-Demenz und Schizophrenie in Verbindung gebracht.
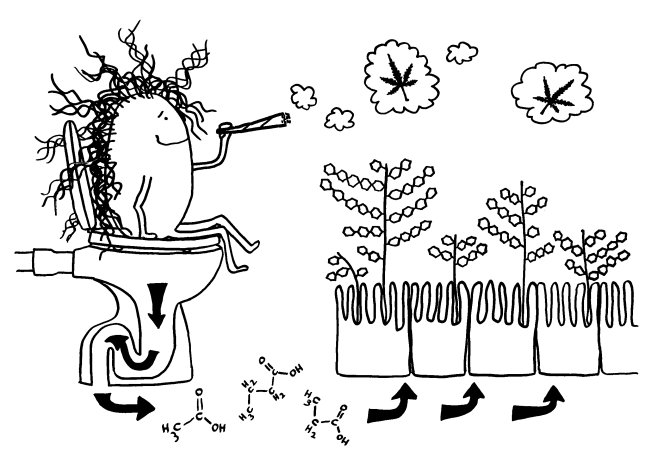
Offenbar wird ihre Aktivität unter anderem von unserer Darmflora reguliert. Vermittelt wird diese Fernwirkung vermutlich über sogenannte kurzkettige Fettsäuren (short-chained fetty acids = SCFA), also Gärungsprodukte wie Essig-, Propion- und Buttersäure, die die Darmbakterien aus unserer Nahrung gewinnen. Diese durchdringen das Darmepithel und gelangen dann entweder selbst über das Blut ins Gehirn, oder sie regen in unserem Darmgewebe Zellen zur Ausschüttung von Botenstoffen an, die dann ihrerseits über die Adern ins Gehirn kommen.
Im Gehirn von Menschen, die ein hohes Schizophrenie-Risiko haben oder sich bereits in der Frühphase der Erkrankung befinden, ist die Konzentration von Zytokinen erhöht; ihre graue Materie geht zurück, und ihre Mikroglia sind überaktiv: Anzeichen für eine Entzündung. Je stärker ihre Mikroglia aktiviert sind, desto stärker sind die Schizophrenie-Symptome, wenn die Erkrankung schließlich ausbricht. Die zeitliche Abfolge lässt vermuten, dass die Mikroglia-Aktivierung nicht lediglich eine Folge einer bereits eingetretenen Störung im Gehirn ist, sondern diese mit verursacht. Dazu passt auch das Lebensalter, in dem Schizophrenie und weitere psychische Erkrankungen besonders häufig ausbrechen: während oder kurz nach der Adoleszenz – genau dann, wenn die Mikroglia im Frontalkortex viel Pruning betreiben.
Literatur:
D. Erny et al.: Host microbiota constantly control maturation and function of microglia in the CNS. Nature Neuroscience 18, 965–977 (2015), doi:10.1038/nn.4030 (nur Abstract frei)
Dazu auch Katrin Zöfel: Bakterien für ein gesundes Gehirn (09.10.2015)
P. S. Bloomfield et al.: Microglial Activity in People at Ultra High Risk of Psychosis and in Schizophrenia: An [11C]PBR28 PET Brain Imaging Study. American Journal of Psychiatry, http://dx.doi.org/10.1176/appi.ajp.2015.14101358 (nur Abstract frei)
Dazu auch Mo Costandi: Brain’s immune cells hyperactive in schizophrenia (16.10.2015)