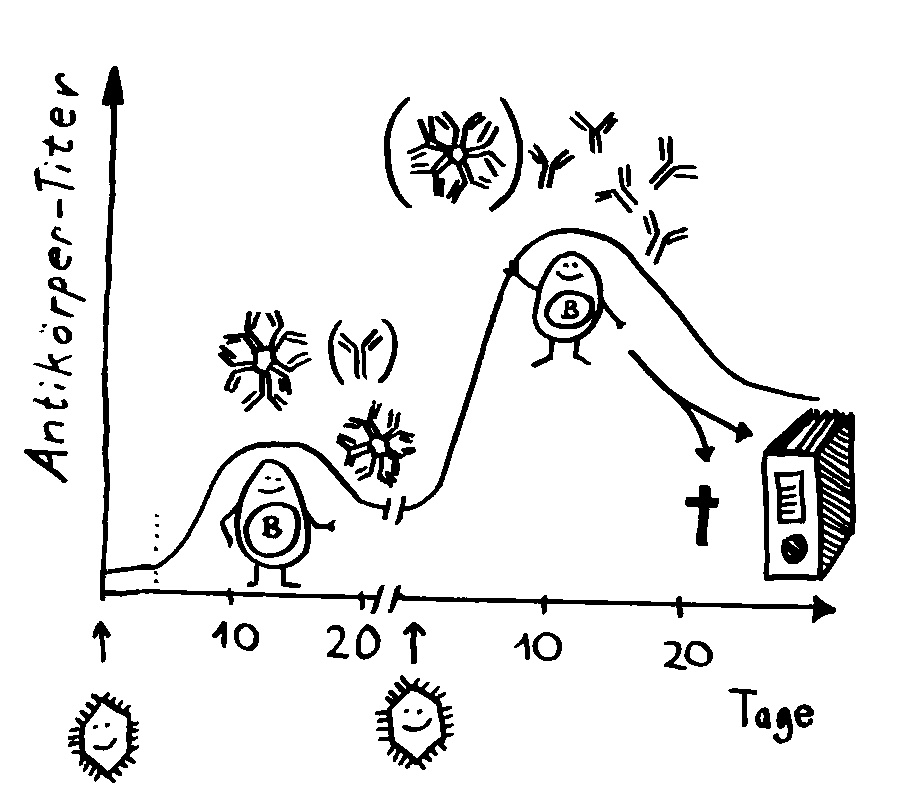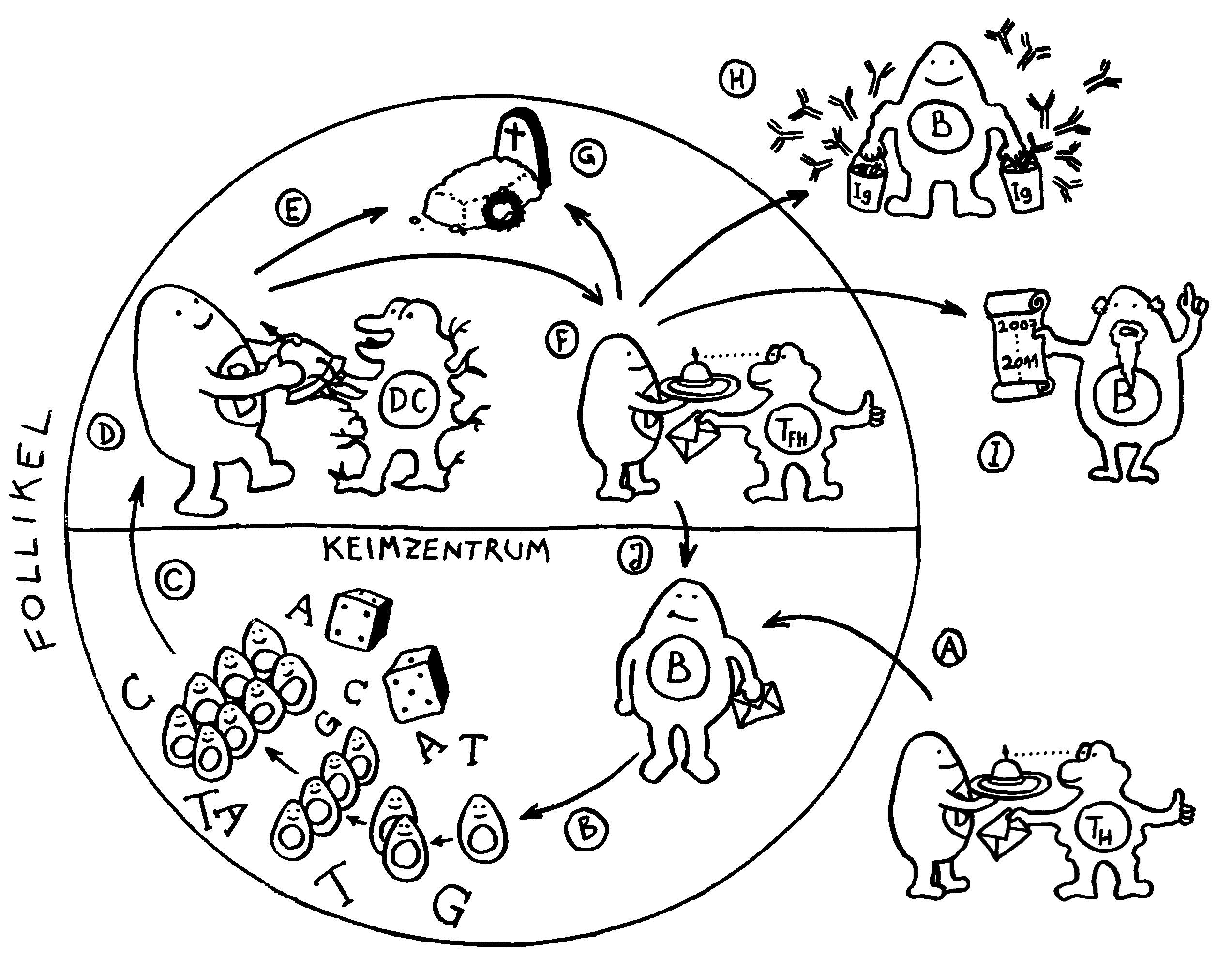
In den Follikeln der Lymphknoten und des tertiären Lymphgewebes kommt es zur Affinitätsreifung. Sie erhöht die Antigen-Bindungsstärke der Immunglobuline, also der B-Zell-Rezeptoren und der Antikörper. Im Uhrzeigersinn, bei 4 Uhr beginnend:
A Eine B-Zelle, die bereits ein Antigen aufgenommen hat, präsentiert ihren Fund einer T-Helferzelle (TH) und wird vollends aktiviert, sofern der T-Zell-Rezeptor das Antigen erkennt. Sie erhält von der T-Helferzelle die Lizenz, in das Keimzentrum des Follikels einzutreten.
B Im Keimzentrum des Follikels vermehrt sich die B-Zelle stark durch Teilung. Währenddessen verändert ein Enzym nach dem Zufallsprinzip einzelne Basen (A, T, C, G) in dem Gen, das die antigenspezifische Bindungsstelle des Immunglobulins codiert. Diesen Vorgang nennt man somatische Hypermutation.
C Die mutierten B-Zellen treten aus der dunklen Zone des Keimzentrums in die helle Zone über, wo sie von dendritischen Zellen (DC) erwartet werden. Diese sind für die Selektion zuständig.
D Die dendritischen Zellen präsentieren ihnen das Antigen, um die Bindungsstärke des mutierten B-Zell-Rezeptors zu prüfen.
E Hat die Mutation die Bindung der Immunglobuline an das Antigen geschwächt, stirbt die B-Zelle durch Apoptose kontrolliert ab.
F Hat die Mutation die spezifische Bindung an das Antigen gestärkt, so führt die B-Zelle dieses Antigen nun auf ihrem MHC-Klasse-II-Molekül einer follikulären T-Helferzelle (TFH) vor, die es mit ihrem spezifischen T-Zell-Rezeptor erkennt. Durch diesen Kontakt wird der Klassenwechsel bei den Immunglobulinen ausgelöst, sodass die B-Zelle nun kein IgM mehr herstellt, sondern IgG, IgE oder IgA – je nachdem, welchen Botenstoff die follikuläre T-Helferzelle ausschüttet. Dazu mehr in der nächsten Abbildung. Je nach dem Ergebnis dieser zweiten Prüfung schlägt die B-Zelle einen von vier Wegen ein:
G Die B-Zelle ist unbrauchbar, weil sie der T-Zelle ihr Antigen nicht effizient präsentiert, und stirbt.
H Die B-Zelle ist zur humoralen Abwehr geeignet, verlässt das Keimzentrum und entwickelt sich zur Plasmazelle weiter, die massenhaft Antikörper erzeugt.
I Einige B-Zellen reifen stattdessen zu Gedächtniszellen heran, die mit ihrem Wissen um die aktuelle Infektion dafür sorgen, dass das Immunsystem auf ein späteres erneutes Auftreten desselben Antigens schneller und stärker reagiert.
J Einige besonders schlagkräftige B-Zellen erhalten die Order, erneut in das Keimzentrum einzutreten, um sich zu vermehren und durch Mutation und Selektion weiter zu verbessern. So steigert der Organismus die Affinität der Immunglobuline zu einem bestimmten Antigen mit der Zeit.
Sie dürfen diese Zeichnung gerne in Folien etc. übernehmen, sofern Sie die Quelle angeben: Dr. Andrea Kamphuis, https://autoimmunbuch.de