
Ich komme noch einmal auf die Meldung zurück, der zufolge unsere Gewebsmakrophagen überwiegend nicht von Stammzellen im Knochenmark abstammen, sondern von Vorläufern aus dem embryonalen Dottersack. Die meisten Nichtbiologen werden das schulterzuckend zur Kenntnis nehmen. Für Biologen ist es eine Sensation, da man bisher davon ausging, dass all unser Zellen, Gewebe und Organe auf eines der drei Keimblätter – Ectoderm, Mesoderm und Endoderm – zurückgehen, die während der sehr frühen Embryonalentwicklung angelegt werden.

Pflanzenkeim mit Keimblättern und erstem echtem Blätterpaar
Die Bezeichnung „Keimblätter“ erleichtert das Verständnis dieser Vorgänge leider überhaupt nicht, denn man denkt bei dem Wort an die ersten Blättchen keimender Pflanzen. Aus diesen simpel gebauten Blättchen entsteht aber später gar nichts; sie schützen und versorgen die junge Pflanze, bis sie die ersten echten Blätter ausgebildet hat, und verkümmern dann.
Das ist bei den dreikeimblättrigen Tieren (allen bilateralsymmetrischen Tieren, also solchen mit einer linken und einer rechten Körperseite) ganz anders – und damit auch bei den Menschen. Während der sogenannten Gastrulation faltet sich der bis dahin zweischichtige Keim zu einem komplexeren dreischichtigen Gebilde zusammen.
Aus der äußeren, oberen Schicht, dem Ektoderm, werden später unter anderem die Haut und das Nervensystem. Aus der inneren, nach konventioneller Darstellung untersten, zum Dottersack orientierten Schicht, dem Endoderm, gehen unter anderem der Verdauungstrakt, die Leber und die Lunge hervor. Und die zuletzt durch Einwanderung von Zellen zwischen die beiden anderen Zelllagen entstandene mittlere Schicht, das Mesoderm, bringt später die Knochen, Muskeln und Nieren, das Herz und das Blut hervor – also auch die Stammzellen im Knochenmark, aus denen unsere Immunzellen entstehen.
Querschnitt durch eine junge Froschlarve mit ersten Gewebetypen und Organen, die aus den drei Keimblättern entstanden sind. Oben das Neuralrohr, in der Mitte der Darm.
Mit jedem Entwicklungsschritt wird das Schicksal der embryonalen Zellen weiter festgelegt: Bestimmte Gene werden abgelesen, andere nicht. Die Zelloberfläche weist entsprechend bestimmte Marker auf und andere nicht. Durch diese Marker zeigen die Zellen ihre Bestimmung an – sowohl ihr örtliches Ziel im Organismus als auch ihre Aufgabe.
Die meisten Zellen prägen ihre endgültige Bestimmung schon früh aus und sind damit festgelegt: Eine Muskelzelle kann sich zum Beispiel niemals in eine Darmschleimhautzelle verwandeln. Nur die sogenannten Stammzellen behalten lange das Potenzial, sich je nach Bedarf in die eine oder die andere Richtung zu entwickeln. Doch auch sie durchlaufen während ihrer Zellteilungen eine Kaskade immer weiterer Festlegungen durch Expression bestimmter Gene: Aus einer sogenannten totipotenten wird zunächst eine pluripotente Stammzelle, aus der noch mehrere, aber nicht mehr alle möglichen Zelltypen heranreifen können. Nach einigen Teilungen verliert die Zelle auch dieses Potenzial und ist dann auf einen Zelltyp – und damit auf dessen Funktionen – festgelegt, was sich wiederum in ihren Oberflächenmarkern ausdrückt.
Manche Immunzellen bewahren sich offenbar lange eine gewisse Restflexibilität. So können T-Helferzellen wohl durch Zellkontakte und Botenstoffe angeregt werden, sich in regulatorische T-Zellen zu verwandeln, die Abwehrreaktionen hemmen, statt sie – wie zuvor – zu verstärken. Aber aus einer B-Zelle kann niemals eine T-Zelle werden und umgekehrt.
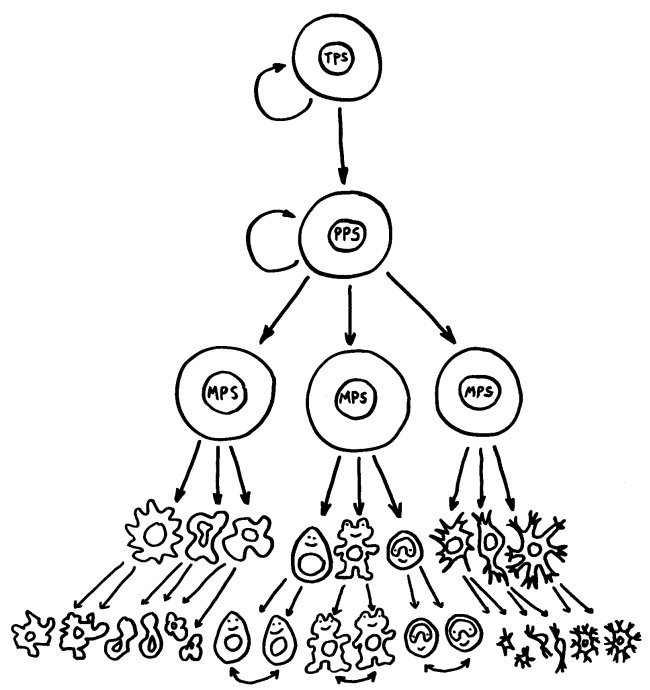
Totipotente, pluripotente und multipotente Stammzellen sowie – ganz unten – fertig ausdifferenzierte Zellen. Eine T-Zelle kann sich u. U. in einen anderen T-Zell-Typ verwandeln, aber nicht in eine B-Zelle oder einen Monozyt (Pfeile ganz unten).
Die Gewebsmakrophagen umgehen den ersten Teil dieser Differenzierungskaskade, indem sie aus dem Dottersack in den Embryo einwandern, während dieser seine Keimblätter bereits voll ausgebildet hat. Verwirrenderweise werden sie mal als embryonale, dann wieder als extraembryonale Zellen bezeichnet. Das ist eine Frage der Perspektive: Sie sind weder rein mütterlichen noch gar rein väterlichen Ursprungs, sondern wie der gesamte Dottersack aus der befruchteten Eizelle, der Zygote, hervorgegangen und insofern embryonal. Andererseits liegt der Dottersack außerhalb des sich entwickelnden Embryos und trägt – mit der nun bekannt gewordenen Ausnahme – keine Zellen zum späteren Organismus bei; insofern ist er extraembryonal.
Dass man die Herkunft der Gewebsmakrophagen erst jetzt aufgedeckt hat, liegt wesentlich an ihrer enormen Ähnlichkeit mit den anderen, aus dem Blut stammenden Makrophagen, die auf dem üblichen Weg während der Hämatopoese aus Vorläufern im Knochenmark entstehen. Sie übernehmen auch ähnliche Aufgaben, fressen nämlich störende oder gefährliche Zellen, Zellreste oder Fremdkörper im Gewebe auf.
Diese Ähnlichkeit trotz ganz anderer Herkunft erinnert mich an die sogenannte konvergente Evolution, also die Entstehung ähnlicher Strukturen auf ganz unterschiedlichen Ästen und Zweigen des Stammbaums des Lebens aufgrund ähnlicher ökologischer Nischen. So gibt (bzw. gab) es in Australien alle möglichen Beuteltiere, die durch den Selektionsdruck in ihrer Umwelt ähnliche Lebensweisen und einen ähnlichen Körperbau hervorgebracht haben wie Säugetiere auf anderen Kontinenten – etwa den Beutelwolf oder die Beutelratte.
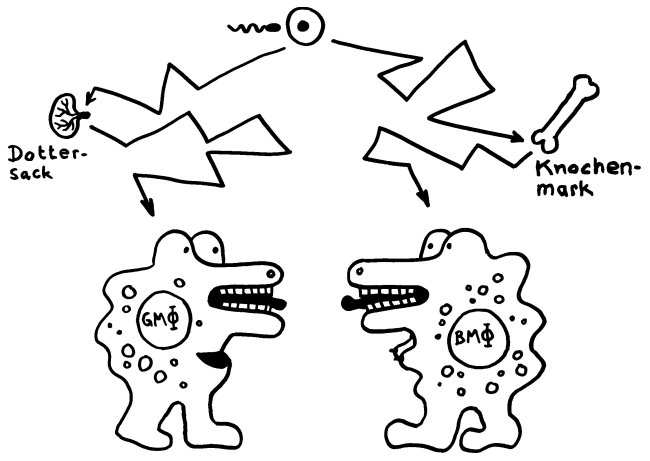
Wie manche Beuteltiere und höhere Säugetiere ähneln sich Gewebsmakrophagen (links) und Blut-Makrophagen (rechts) sehr, obwohl sie ganz unterschiedlicher Abstammung sind: Die einen gehen auf den Dottersack zurück, die anderen auf das Knochenmark.
Und so, wie beide konvergent entstandene Formen nur zeitgleich existieren können, weil sie räumlich getrennt sind, so haben auch Gewebsmakrophagen und „richtige“ Makrophagen normalerweise unterschiedliche Lebensräume, nämlich einerseits die Gewebe und andererseits Blut und Lymphe. Nur bei einer Entzündung dringen die „richtigen“ Makrophagen kurzfristig in das Revier der Gewebsmakrophagen ein, um dort zu helfen.