

Im vorigen Beitrag habe ich den Unterschied zwischen den Reproduktionsstrategien von Maus und Mensch erwähnt, der – neben anderen guten Gründen – die Übertragbarkeit von Ergebnissen immunologischer Studien an Labormäusen auf Menschen erschwert. Das will ich hier genauer ausführen.
Theoretische Ökologie, erste Lektion: Malthus’sche Gleichung. Der Mathematiker Thomas Malthus beschrieb 1798 in seinem Essay on the Principle of Population das Wachstum einer Bevölkerung, deren Geburtenrate über der Sterberate liegt. Eine anfangs kleine Gründerpopulation in einem neuen Lebensraum wächst zunächst exponentiell an, und zwar mit der Wachstumsrate r: der Differenz aus Geburten- und Sterberate.
Zweite Lektion: die logistische Gleichung, eingeführt 1837 vom Mathematiker Pierre François Verhulst. Wenn die verfügbaren Ressourcen nicht ebenfalls exponentiell anwachsen, schwächt sich der Zuwachs durch Konkurrenz um diese knappen Ressourcen ab – und zwar umso stärker, je näher die Populationsgröße N der sogenannten Kapazitätsgrenze K kommt, der maximalen Tragfähigkeit der Umwelt. Die Populationsentwicklung nimmt eine S-Kurve; in einer stabilen Umwelt halten sich schließlich Zuwachs und Schwund die Waage.
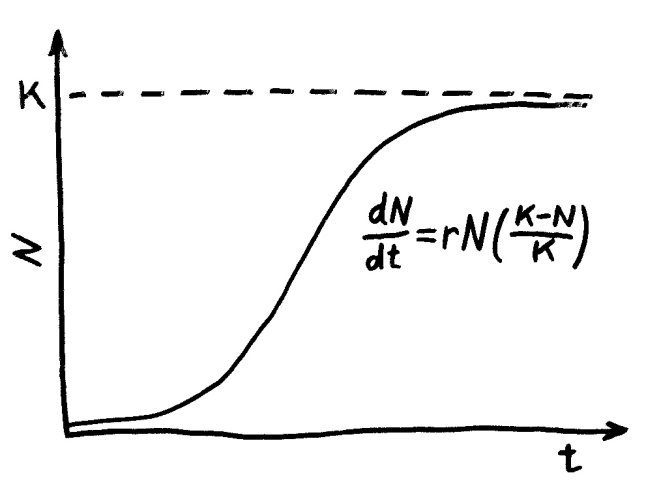
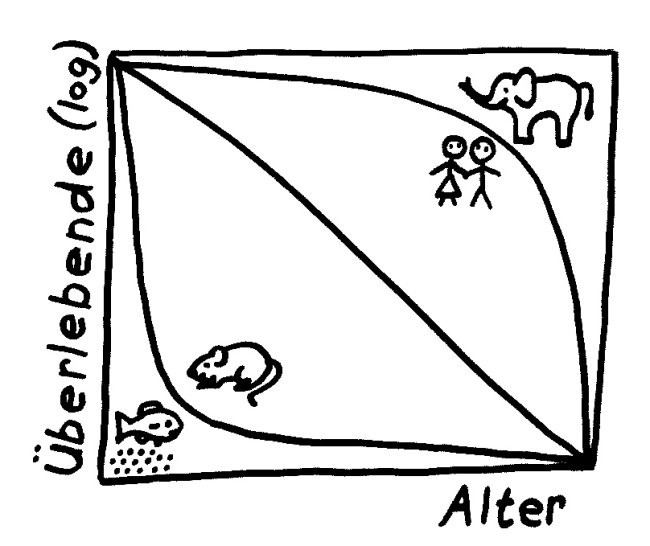
r- und K-Strategen
Je nach Lebensraum, Lebensweise, Lebenserwartung usw. bilden sich bei Tierarten evolutionär unterschiedliche Reproduktionsstrategien heraus. Die beiden Extreme werden nach den eben eingeführten Größen r und K als r-Strategie und K-Strategie bezeichnet. Typische r-Strategen wie Insekten oder Fische setzen enorm viel Nachwuchs in die Welt, in den sie dann wenig investieren, sodass ein Großteil bald zugrunde geht und nur wenige Exemplare die Geschlechtsreife erreichen. Typische K-Strategen wie Elefanten haben wenig Nachwuchs. Dafür kümmern sie sich intensiv und lange um diese wenigen Kinder, sodass viele von ihnen die Geschlechtsreife erreichen – und darüber hinaus ein hohes Alter, denn sie müssen sich ja ihrerseits lange um ihren Nachwuchs kümmern.
Auch innerhalb der Klasse der Säugetiere ist das Spektrum breit. Mäuse und Ratten sind r-Strategen, Menschen K-Strategen. Sie haben eine ganz unterschiedliche life history – ein evolutionstheoretischer Fachbegriff, der mit „Lebenslauf“ oder „Lebensgeschichte“ nur unzureichend übersetzt ist: Sie setzen die mühsam der Umwelt und der Konkurrenz abgetrotzten Ressourcen je nach Lebenszyklusphase und Geschlecht unterschiedlich ein.
Wer kurz lebt und so schnell wie möglich so viel Nachwuchs wie möglich zeugen muss, um seine Gene effektiver in die nächste Generation zu tragen als die Konkurrenz, der fängt keine Riesterrente an und geht auch nicht zur Vorsorge. Stattdessen fließen die Mittel in die Ausbildung primärer, sekundärer und tertiärer Geschlechtsmerkmale: schnelle Spermien, stramme Muskeln, Rauflust und so weiter. Wer zwanzig Jahre braucht, bis er ein Kind bekommt, und noch einmal zwanzig, um es großzuziehen, investiert mehr in die eigene Gesundheit – etwa in ein schlagkräftiges Immunsystem, das Krankheitserreger in Schach hält.
Hormongesteuert
Wie teilt ein Organismus seine Ressourcen ein? Woher „weiß“ er, in welcher Situation und Lebensphase er sie auf welches Konto einzahlen muss? Hier kommen die Hormone ins Spiel, vor allem die Sexualhormone: Unser endokrines System integriert Umweltreize und Körpersignale und erreicht noch den letzten Winkel des Körpers, um dort – etwa über die Steuerung der Erbgut-Ablesung in den Zellen – die nötigen Veränderungen in Gang zu setzen. Gerade zwischen den kostspieligen Systemen Sexualität und Immunabwehr kommt es dabei regelmäßig zu Zielkonflikten (sogenannten trade-offs).
Am besten belegt ist das für Testosteron, das männliche Sexualhormon, das in geringerem Maße auch von Weibchen bzw. Frauen hergestellt und benötigt wird. Es fördert direkt (z. B. durch die Ausbildung körperlicher Geschlechtsmerkmale) und indirekt (etwa durch Förderung von Verhaltensweisen, die Konkurrenten in Schach halten) die Fortpflanzungschancen von Männchen bzw. Männern. Trotz dieses Selektionsdrucks kann seine Produktion nicht in den Himmel wachsen, weil es Immunreaktionen (etwa die Antikörperproduktion) dämpft und damit die Widerstandskraft gegen Krankheitserreger schwächen kann – und zwar bei beiden Geschlechtern.
Umgekehrt wird die Testosteron-Produktion im Lauf der Evolution auch nicht zugunsten eines schlagkräftigeren Immunsystems zurückgefahren, weil das dem Reproduktionsinteresse der Männchen zuwiderliefe – und weil das Immunsystem ohne das immunsuppressive Testosteron ständig übers Ziel hinausschösse, wie es bei Autoimmunstörungen und Sepsen passiert. Durch das Zusammenspiel von sexueller Selektion und natürlicher Selektion kommen weitere Effekte hinzu, die der Intuition zuwiderlaufen können. So kann ein Männchen gerade dadurch ein attraktiver Partner sein, dass es trotz hohem Pathogendruck in der Umwelt und schwachem Immunsystem augenscheinlich gesund ist, also offenbar Verletzungen und Infektionen geschickt aus dem Wege gehen kann.
Die testosteronvermittelten Zielkonflikte zwischen Reproduktion und Selbsterhalt gehen bei r-Strategen viel stärker auf Kosten des Immunsystems als bei K-Strategen, und die Unterschiede zwischen weiblichen und männlichen evolutionären Interessen fallen bei kurzlebigen Tieren stärker aus als bei langlebigen, bei polygamen Arten stärker als bei monogamen Arten und bei Arten ohne männliche Beteiligung an der Aufzucht stärker als bei solchen, in denen sich beide Geschlechter um die Jungen kümmern. Sie spielen folglich bei Hausmäusen eine größere Rolle als bei Menschen – auch wenn beide Arten nicht an den äußersten Enden der Spektren stehen. (So richtig monogam sind wir zum Beispiel nicht.)
Labormäuse …
Warum sind Mäuse dennoch so beliebte Labortiere, auch in der Immunologie? Eben drum: weil sie schnell geschlechtsreif werden und viele Junge kriegen. Weil sie nicht viel kosten. Und weil wir wegen der evolutionären Distanz und ihrer Rolle als Vorratsschädlinge wenig Hemmungen haben, mit ihnen Experimente anzustellen, die wir an Menschen niemals durchführen könnten.
Die Labormaus stammt von der Hausmaus (Mus musculus) ab, von der es eine westliche (Mus musculus domesticus) und eine östliche Unterart gibt (Mus musculus musculus). Seit den 1920er-Jahren wurden Mäusestämme für die Forschung gezüchtet, und zwar nicht direkt aus Wildfängen, sondern auf der Basis sogenannter Farbmäuse, die schon länger für die Haltung als Haustiere kultiviert wurden. Spätere Analysen haben gezeigt, dass das Genom der Labormaus ein Mosaik aus dem Erbgut beider Unterarten und vielleicht sogar weiterer Zutaten ist.
Durch fortgesetzte Selektion auf eine hohe Reproduktionsrate sind weibliche Labormäuse heute sehr gute Futterverwerter und damit etwa drei Wochen früher geschlechtsreif als ihre wild lebenden Artgenossinnen: Die Hälfte der Labortiere ist mit 40 Tagen so weit, die Hälfte der Wildtiere dagegen erst mit 60 bis 70 Tagen. Bei den Männchen hat sich die Geschlechtsreife nicht vorverlagert; sie benötigen nach wie vor knapp 40 Tage. Nach drei Wochen Tragzeit kommen pro Wurf bis zu 12 nackte Nestlinge zur Welt; ein Weibchen schafft theoretisch bis zu acht Würfe im Jahr. Seine Lebenserwartung beträgt etwa zwei Jahre; die Männchen halten etwas länger durch.
… und ihre wilden Vettern
Wilde Hausmäuse leben entweder in stabilen, dichten Populationen in der Nähe menschlicher Siedlungen, etwa in Scheunen, oder weit verstreut in freier Natur. Dann sind die Populationen instabil, und die Sterblichkeit ist sehr hoch (monatlich etwa 30 Prozent, über den Winter kommen nur 10 Prozent). Anders als im Labor leben die Weibchen im Schnitt länger als die Männchen – vor allem wohl, weil sie nicht so viele Würfe haben und weil die Männchen größere Risiken eingehen als in der Käfighaltung.
Angesichts der immensen Verbreitung von Labormäusen und der weit reichenden Schlüsse, die aus medizinischen, insbesondere immunologischen und onkologischen Studien an diesen Tieren gezogen werden, gibt es bestürzend wenige Untersuchungen über das Immunsystem ihrer wilden Vettern: genau drei. Das Immunsystem von Mäusen aus Wildfängen reagiert demnach auf gezielt verabreichte Antigene stärker als das Immunsystem ihrer zahmen Vettern. In ihren peripheren Lymphknoten finden sich zum Beispiel voraktivierte natürliche Killerzellen, die zudem stärker auf die Stimulation mit Zytokinen ansprechen. Außerdem variiert die Heftigkeit der Immunreaktionen zwischen den wilden Individuen stärker als zwischen Labortieren.
Beides ist mit der individuellen immunologischen Vorgeschichte zu erklären, also mit dem Spektrum der Antigene, denen die Tiere in der Natur ausgesetzt waren und die sie erfolgreich bekämpft haben. Um die Immunreaktionen von Menschen und Mäusen vergleichen zu können, sollte die Forschung viel mehr mit Wildfang-Mäusen arbeiten. Denn Menschen leben nun einmal nicht in sterilen Welten; ihr Immunsystem ist ebenfalls an zahlreichen Infektionen, Verletzungen und anderen Ereignissen gereift. Doch selbst dann wären die Ergebnisse von Mäusestudien nur mit größter Vorsicht auf Menschen übertragbar – wegen der oben erläuterten Unterschiede.
Andere Nager, andere Sitten
Auch die Untersuchung weiterer wilder Nagetiere, etwa der Feldmäuse, kann für Überraschungen sorgen. So ging man aufgrund von Studien an Labormäusen lange davon aus, dass Antioxidantien wie Vitamin C oder Vitamin E, dauerhaft der Nahrung zugesetzt, die verheerende Wirkung von reaktiven Sauerstoff-Spezies (den bösen „freien Radikalen“) in unseren Zellen unterbinden, damit den Alterungsprozess verlangsamen und letztlich das Leben verlängern.
Dann hat man dasselbe mit Erdmäusen probiert, einer Art aus der Gattung der Feldmäuse. Siehe da: Erdmäuse, die regelmäßig Vitamin C oder Vitamin E erhielten, starben früher als die Tiere in den Kontrollgruppen – und zwar dramatisch früher (beispielsweise im Median nach 353 statt nach 477 Tagen). Die Gründe sind noch unklar, und die Nahrungsergänzungsmittel-Industrie wird wenig Interesse daran haben, sie aufzuklären.
Labormäuse als Modelle: Genetisches Geschlecht und Sexualhormone entkoppeln
Das heißt nun nicht, dass Studien an Labormäusen überflüssig oder grundsätzlich irreführend wären. Man muss sich nur ständig bewusst machen, dass Labormäuse Modelle für die Verhältnisse im Menschen sind – mit allen Vor- und Nachteilen. Am faszinierendsten finde ich Versuche, in denen Grundlagenfragen geklärt werden, die beim Menschen einfach nicht zu lösen sind: etwa die Frage, ob das häufigere Auftreten von Autoimmunerkrankungen bei Frauen auf die Sexualhormone (insbesondere Estrogen und Testosteron) oder auf die unterschiedliche Ausstattung mit Geschlechtschromosomen (XX bei der Frau, XY beim Mann) zurückzuführen ist. Dazu mehr im nächsten Artikel.
Sie sollten sich die Brutpflege von Mäusen u. Ratten nochmal genauer ansehen und ihr soziales Interagieren miteinander.. wenn diese Tiere r-Strategen sind, sind wir es auch. Ich denke hier spielt die schlichte Größe die tragende Rolle bei der Wurfanzahl. Es lässt sich jedoch nicht klar auf die Prinzipien der Population abschieben.
Ich hatte als Kind eine Maus, die ganz wunderbar mit einer anderen Maus sozial interagierte, und später eine Ratte. Dass sich die Mütter um ihren Wurf kümmern, ändert nichts daran, dass Mäuse und Ratten r-Strategen sind, also vergleichsweise wenig Ressourcen in das einzelne Junge investieren. Das habe ich mir auch nicht ausgedacht, sondern der Fachliteratur entnommen. Die Größe der Tiere spielt eine gewisse Rolle: Ein sehr großes Säugetier kann per se kein r-Stratege sein, weil es lange dauert, bis der Nachwuchs hinreichend groß ist, um allein zurecht zu kommen. Aber der Umkehrschluss gilt nicht: Es gibt kleine Tiere, die K-Strategen sind. Fledermäuse zum Beispiel können ziemlich alt werden und bekommen wenig Junge auf einmal, obwohl sie nicht größer sind als Mäuse und Ratten.