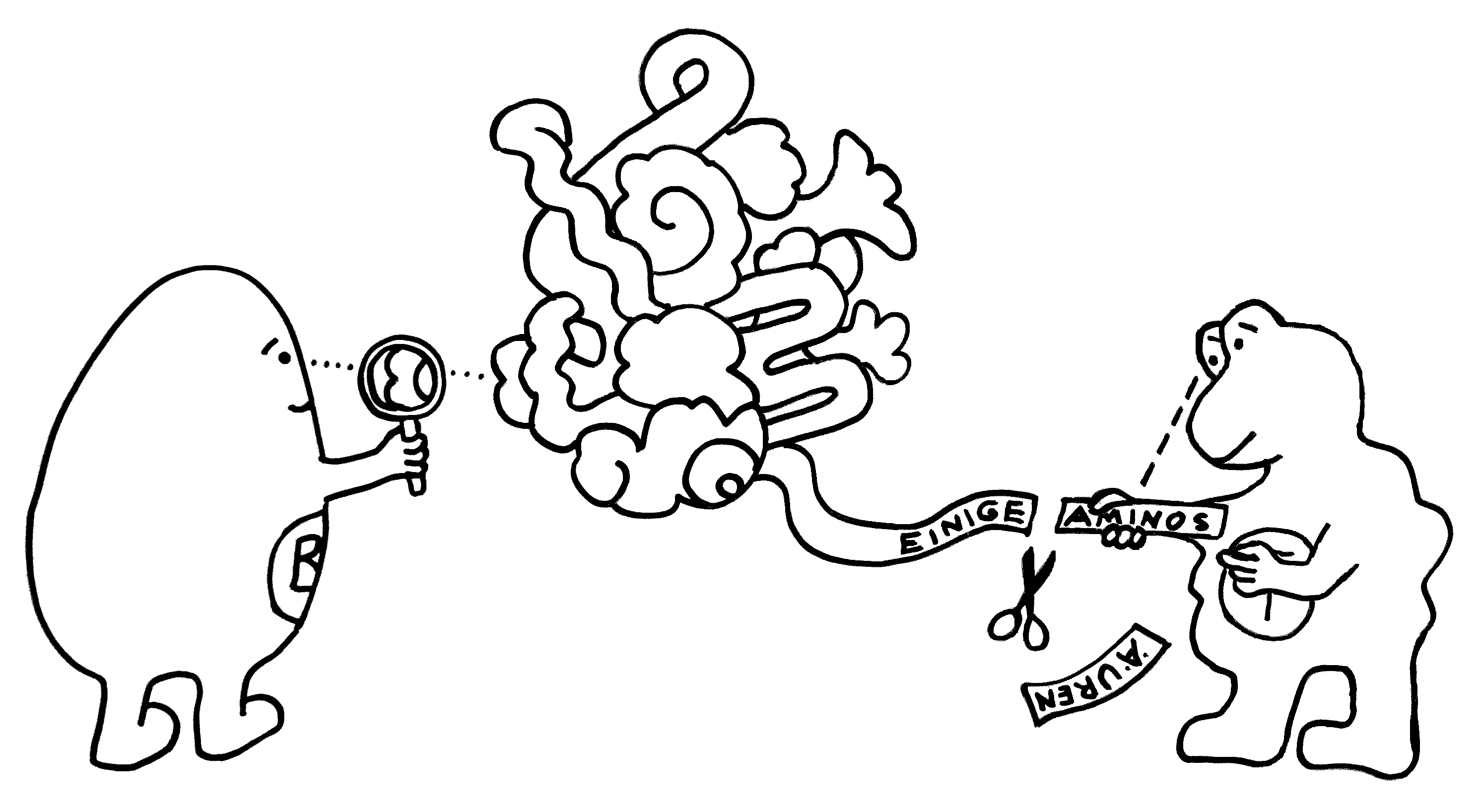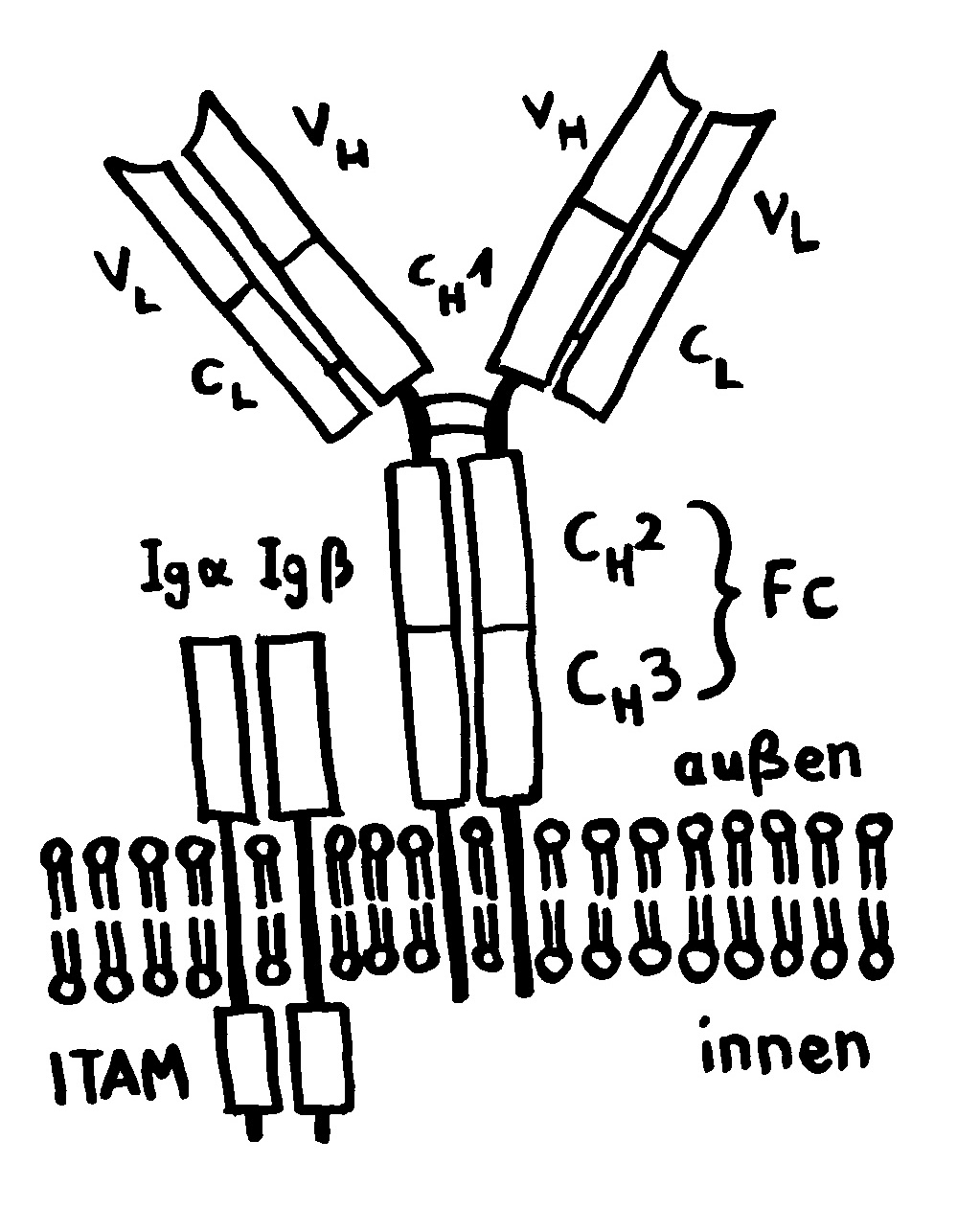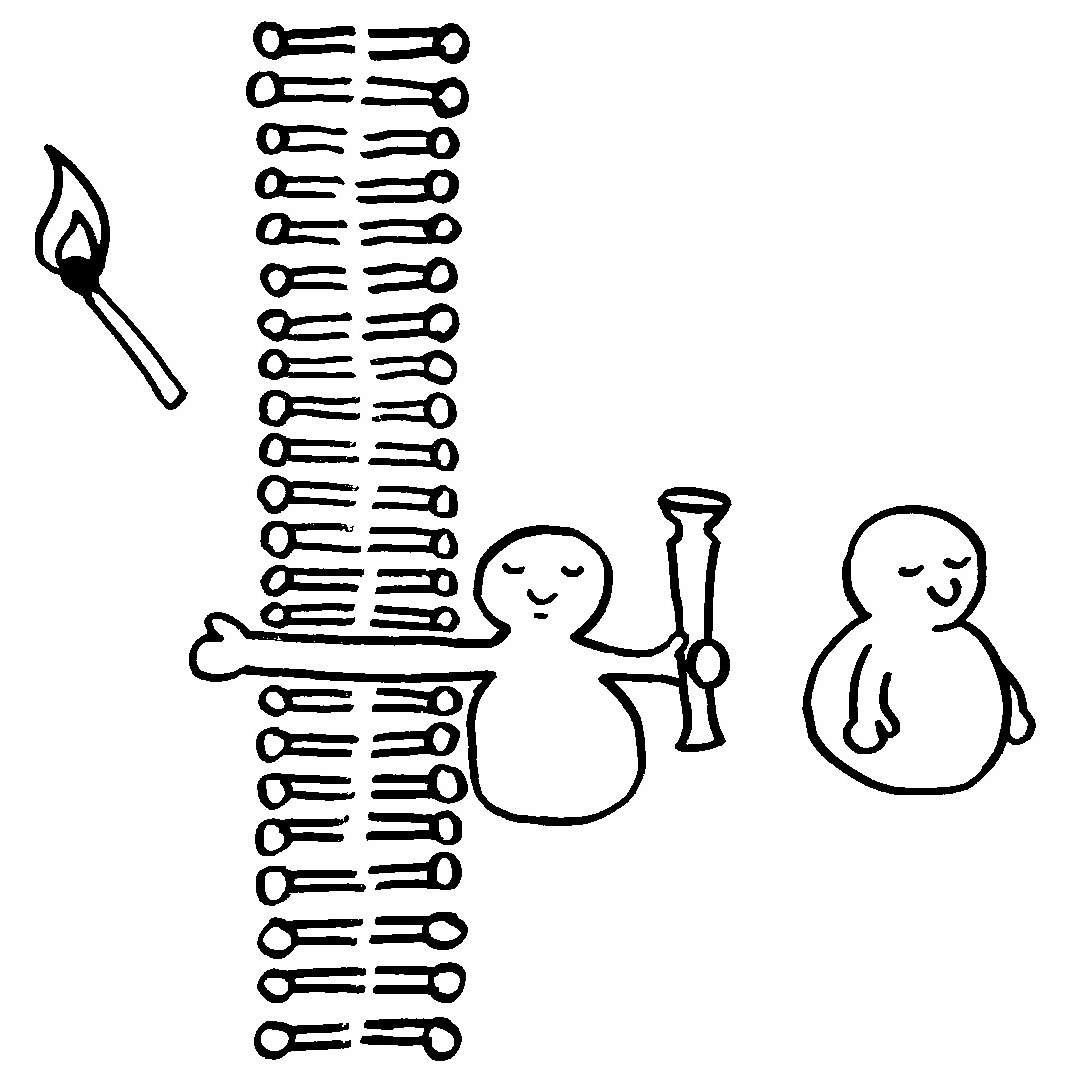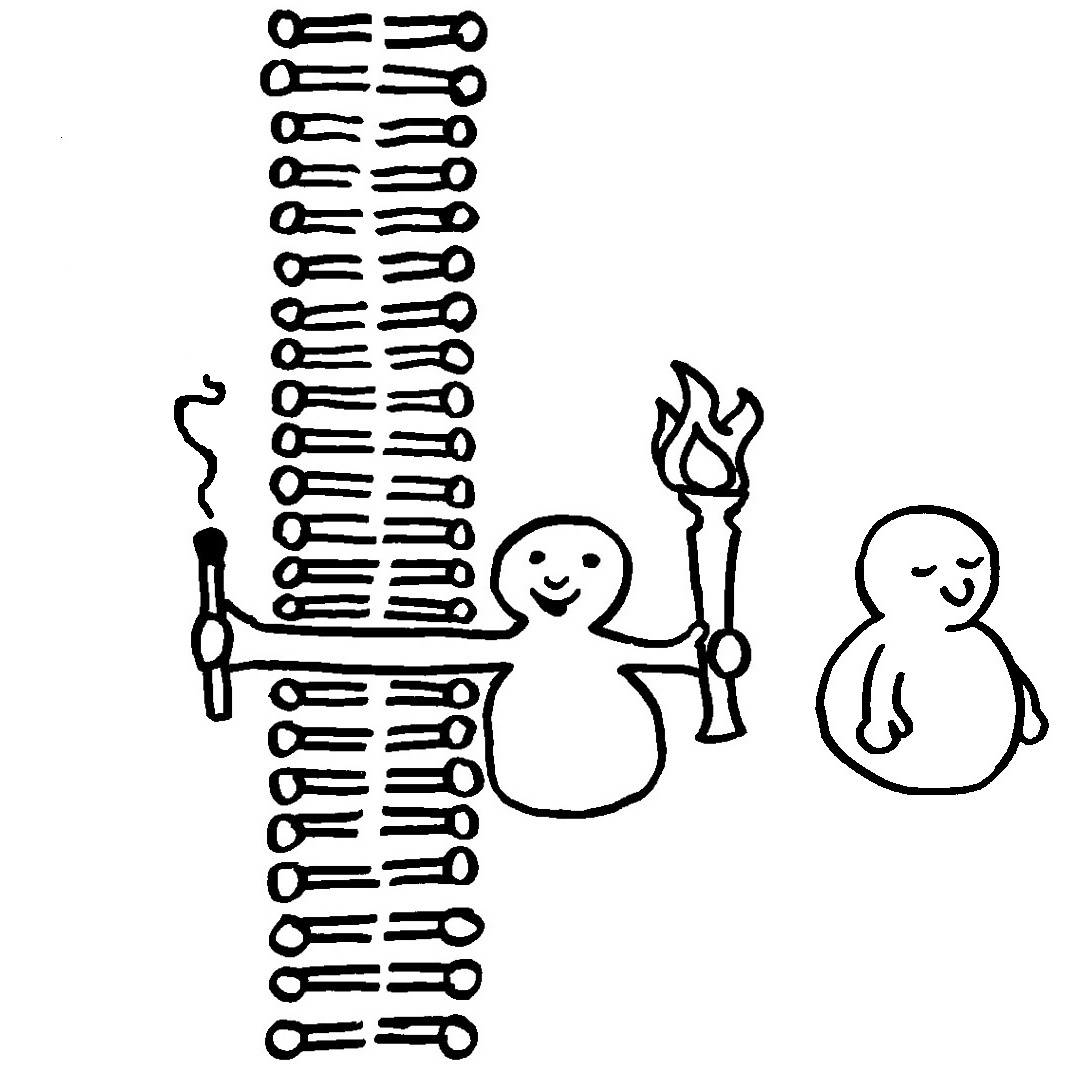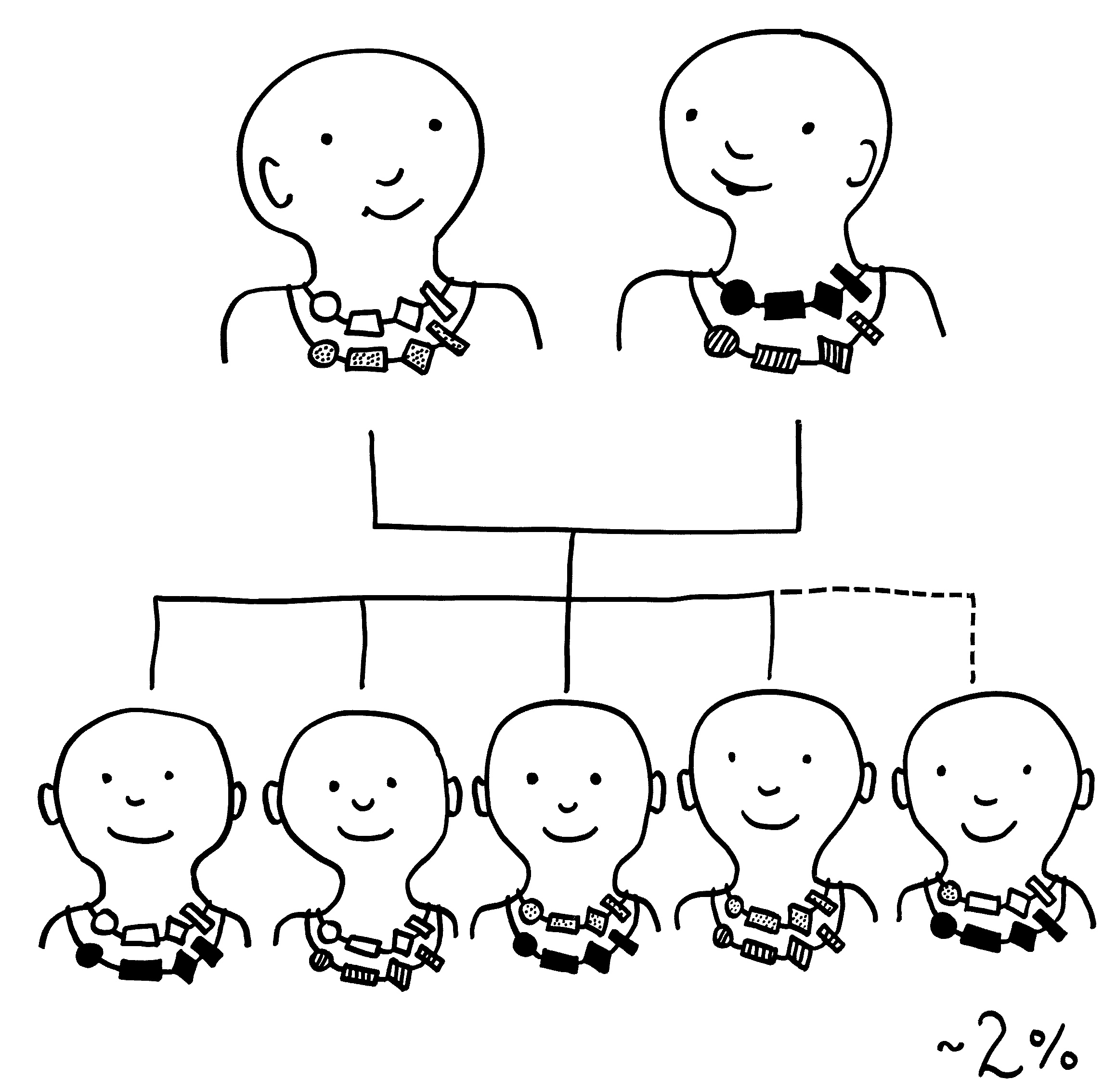
Die meisten MHC-Gene liegen auf unserem Chromosom 6 so eng nebeneinander, dass sie gemeinsam vererbt werden. Die ganze Serie aus MHC-Genvarianten wird als Haplotyp bezeichnet. Jeder Mensch hat zwei Chromosomensätze und damit zwei Haplotypen, je einen auf einem der beiden Chromosomen – hier als zwei Ketten mit je vier unterschiedlichen Perlen (den einzelnen MHC-Genen) dargestellt.
Bei der Fortpflanzung erbt jedes Kind einen väterlichen und einen mütterlichen Haplotyp, was vier gleich wahrscheinliche Kombinationen ergibt. Nur etwa zwei Prozent aller Kinder haben einen veränderten Haplotyp, weil bei der Entstehung der Keimzellen eines Elternteils dessen beide Haplotypen durch Rekombination überkreuzt wurden (Crossing-over, s. Abb. 84 ).
Sie dürfen diese Zeichnung gerne in Folien etc. übernehmen, sofern Sie die Quelle angeben: Dr. Andrea Kamphuis, https://autoimmunbuch.de