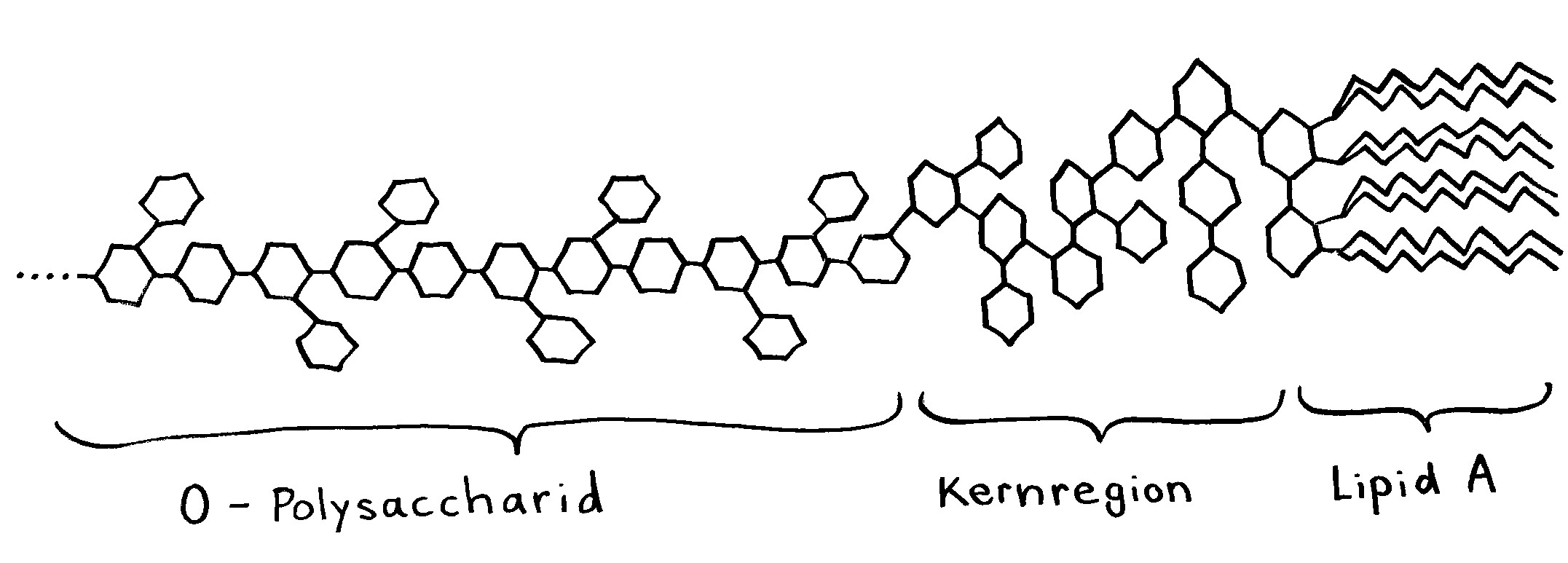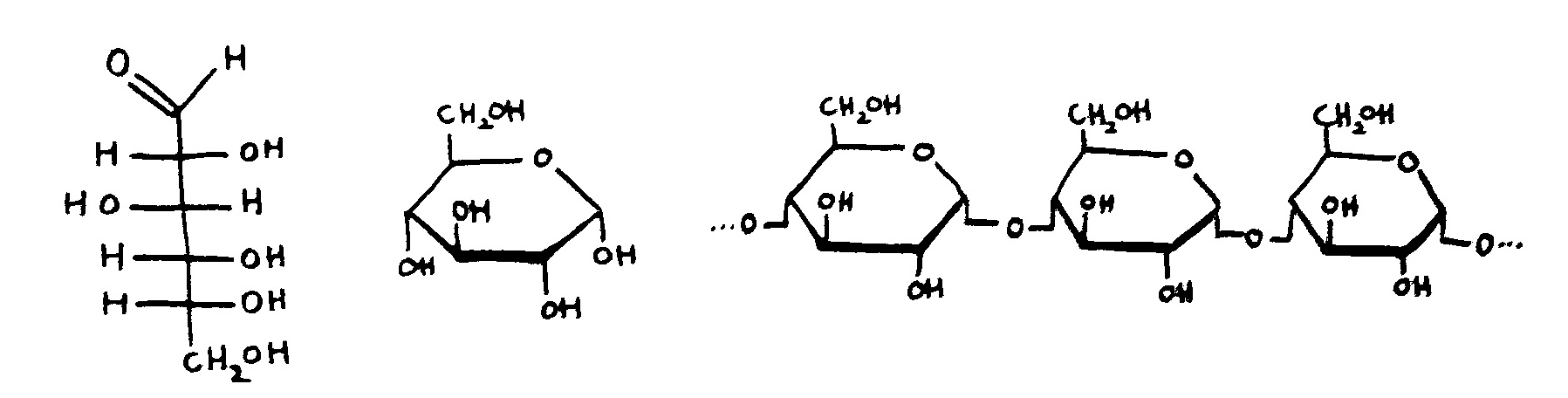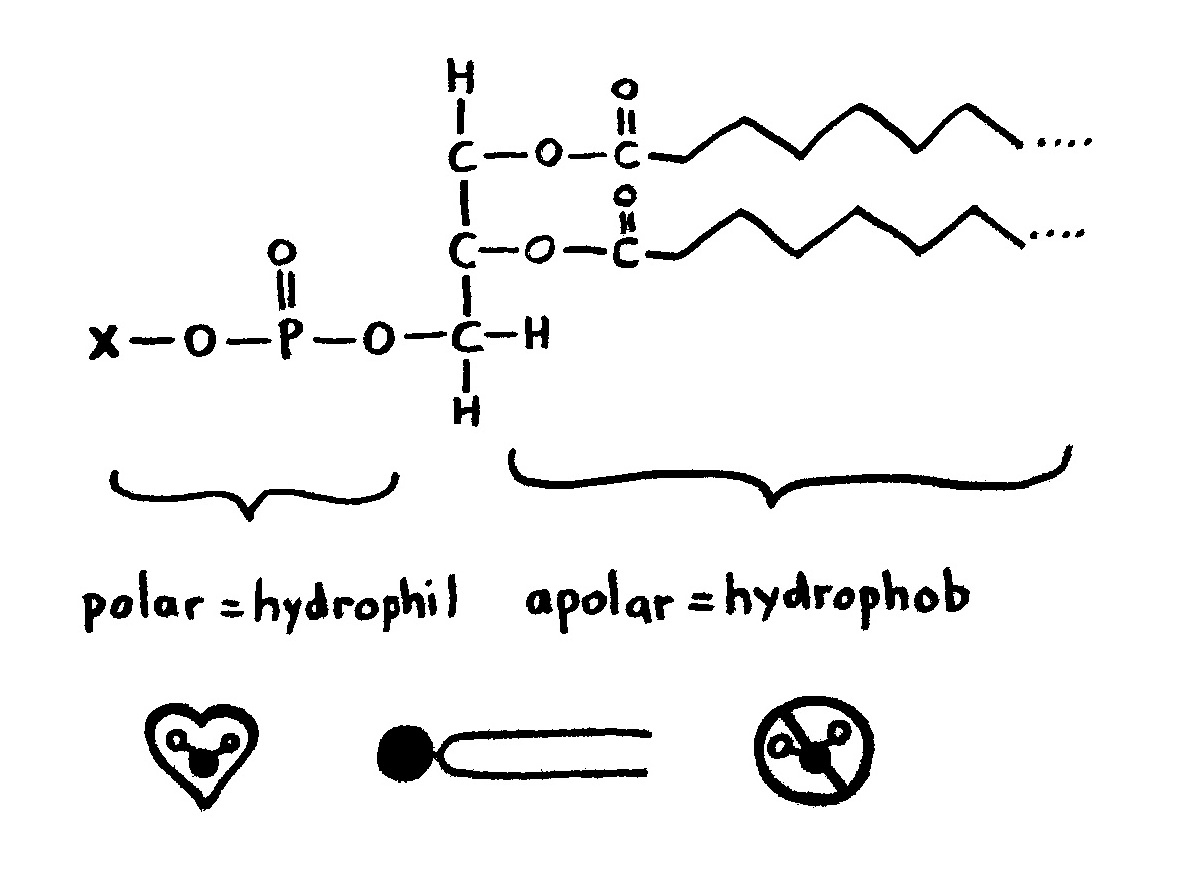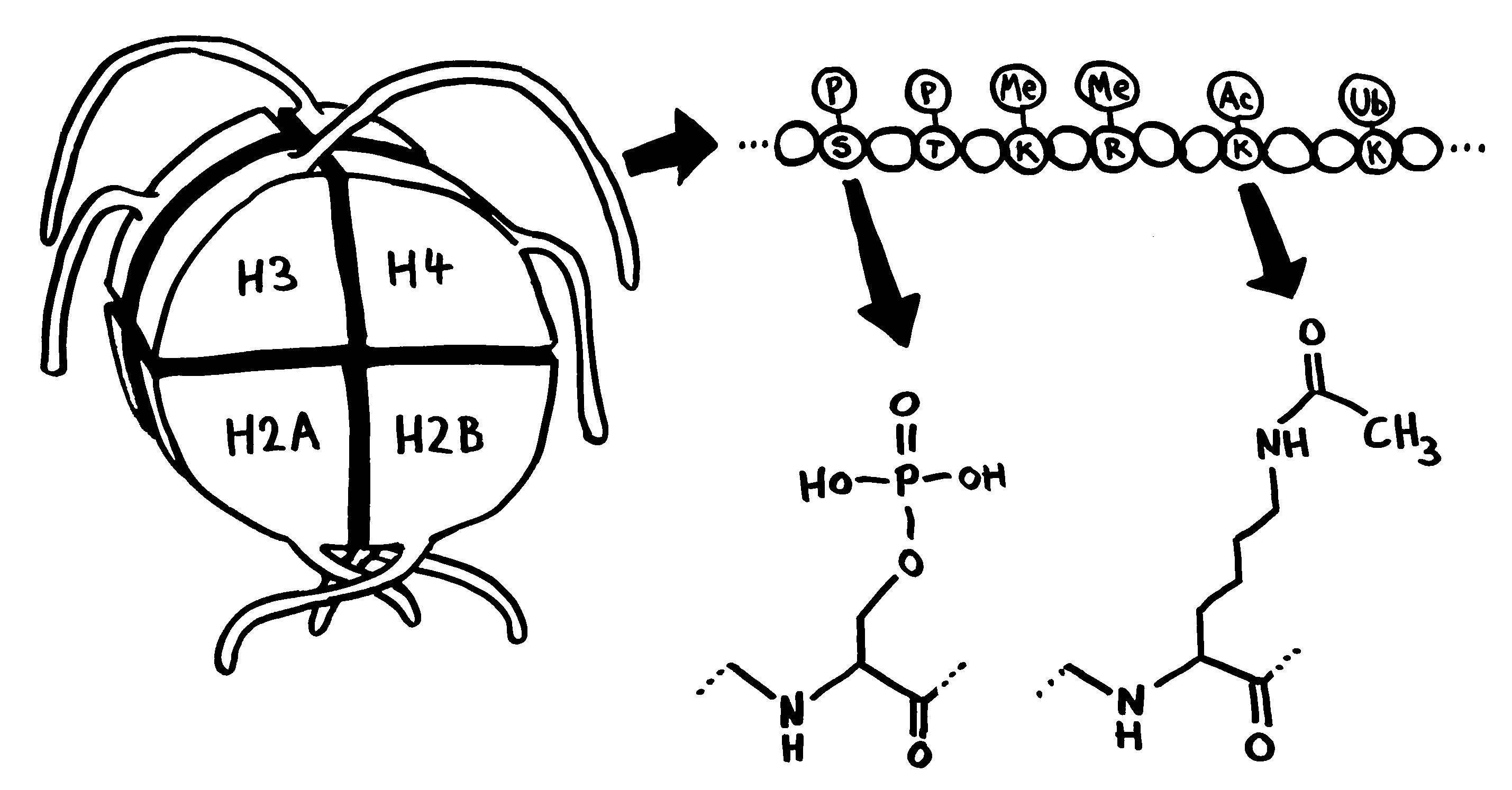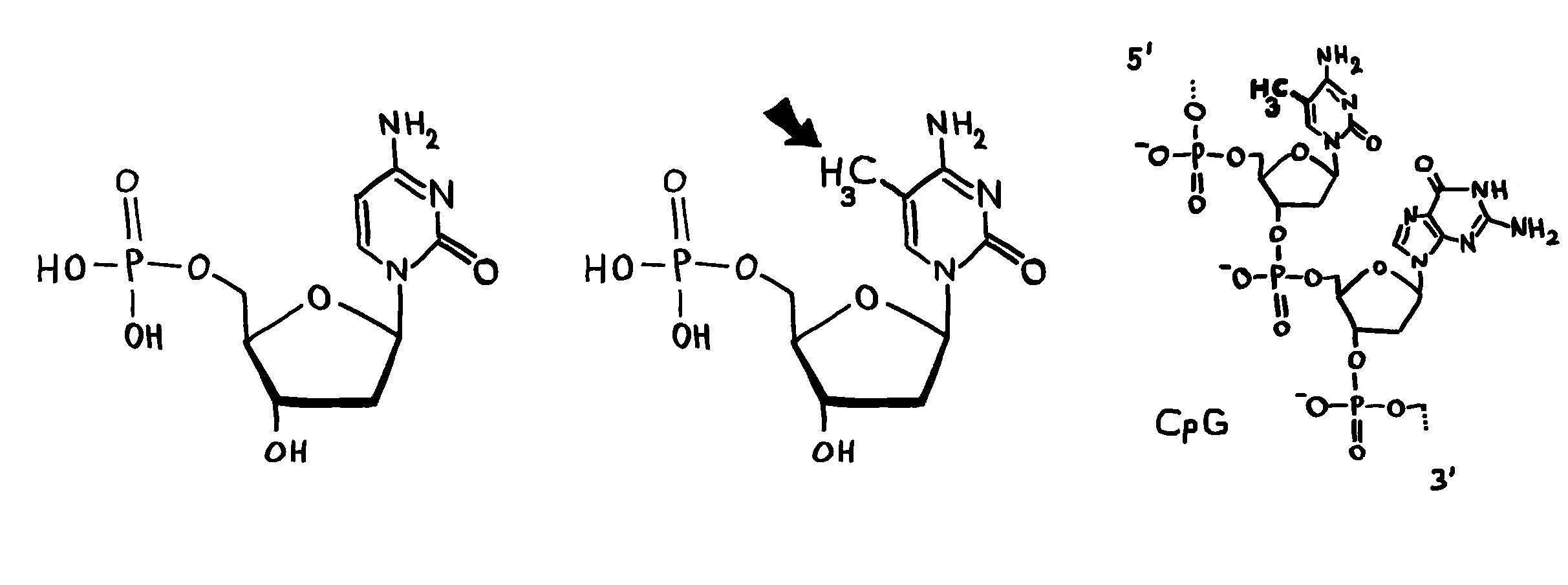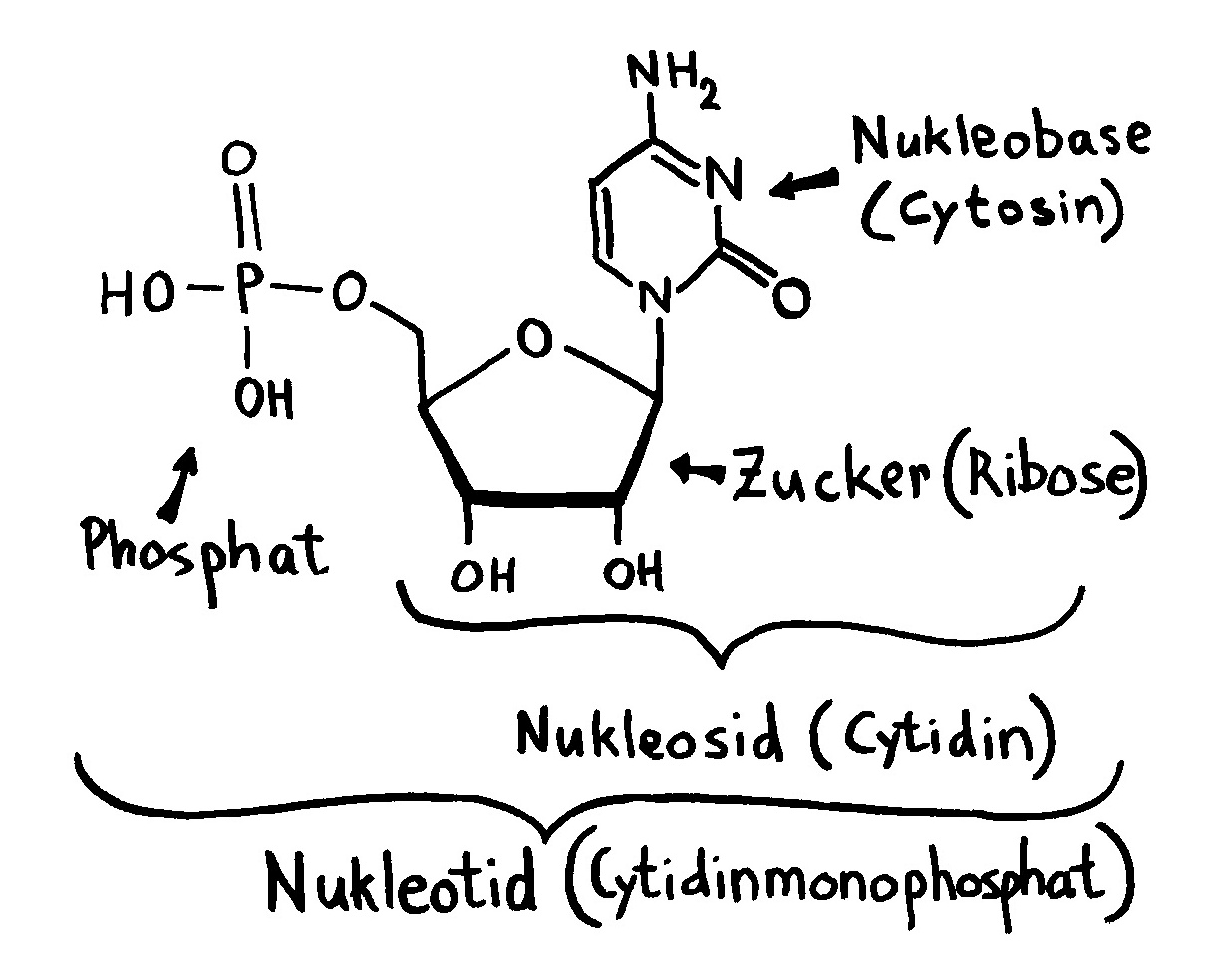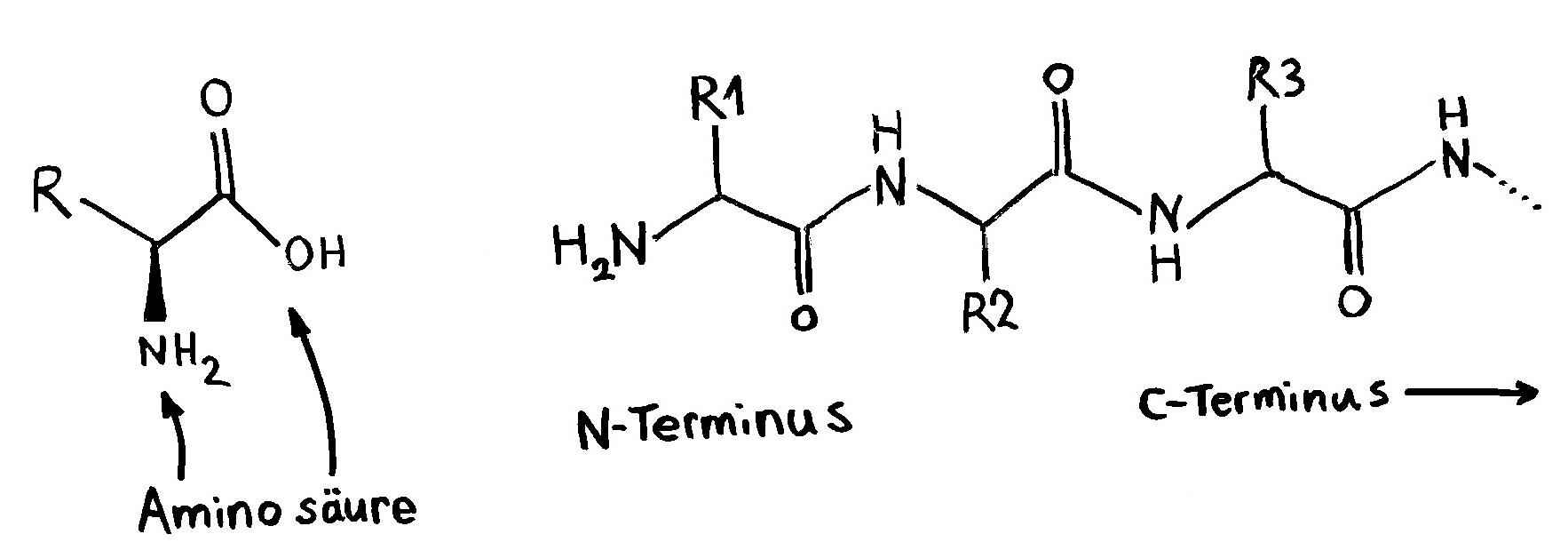
Links: Jede Aminosäure hat eine Amino- und eine Säuregruppe. Die 20 üblicherweise in Proteinen vorkommenden Aminosäuren unterscheiden sich im Rest (R). Rechts: Durch Verbindung der Säuregruppe einer Aminosäure mit der Aminogruppe der nächsten Aminosäure entstehen unter Wasserabscheidung Peptide wie dieses. Wie die Nukleinsäurestränge haben auch Peptide zwei unterschiedliche Enden: den N-Terminus,
der mit einem Stickstoffatom (N) endet, und den C-Terminus, an dem das Kohlenstoffatom der letzten Säuregruppe steht. Peptide winden sich zu dreidimensionalen Gebilden zusammen, deren Form durch die Abfolge der Aminosäuren festgelegt wird.
Sie dürfen diese Zeichnung gerne in Folien etc. übernehmen, sofern Sie die Quelle angeben: Dr. Andrea Kamphuis, https://autoimmunbuch.de