Etliche Bakterien und Viren entziehen sich der Abwehr, indem sie sich ausgerechnet im Inneren von Immunzellen einnisten. Eines der bekanntesten Beispiele ist das Humane Immundefizienz-Virus (HIV), das in T-Zellen überdauert. Viel häufiger und zum Glück weniger gefährlich ist das Epstein-Barr-Virus (EBV), das zu den Herpes-Viren gehört und sich in unseren B-Zellen versteckt. Einmal infiziert, trägt man es so ein Leben lang mit sich herum, und meistens bemerkt man davon nichts.
Bis zum 35. Lebensjahr haben sich über 95 Prozent aller Menschen das Virus zugezogen. Während sich in den Entwicklungsländern – wie früher auch bei uns – die meisten bereits als Kleinkinder symptomfrei anstecken, infizieren sich etliche Menschen in hoch entwickelten Ländern mit guter Hygiene erst als Jugendliche oder junge Erwachsene und entwickeln dann das Pfeiffer-Drüsenfieber. Nach einer akuten Infektionsphase in den Mandeln startet das Virus ein Latenzprogramm: Es nistet sich in langlebigen B-Gedächtniszellen ein, in denen es nicht weiter stört, aber die Funktion der B-Zellen subtil beeinflussen kann. Die B-Gedächtniszellen wandern über die Blutbahn in andere Organe.
Schon lange steht das Virus im Verdacht, bei Menschen mit entsprechender genetischer Veranlagung den Ausbruch von Autoimmunerkrankungen zu fördern, etwa Lupus, Multiple Sklerose, rheumatoide Arthritis, Hashimoto-Thyreoiditis, Sjögren-Syndrom, Typ-1-Diabetes, systemische Sklerose oder chronisch-entzündliche Darmerkrankungen. Gegen welches Organ oder Gewebe sich die Autoimmunreaktionen richten, scheint von ererbten Risikoallelen abzuhängen, insbesondere von bestimmten MHC-Klasse-II-Genvarianten und einer Veranlagung zu einem Mangel an regulatorischen T-Zellen (Tregs). Aber wie tragen die Viren zum Ausbruch der Autoimmunerkrankung bei? Alle möglichen Mechanismen werden in der Literatur diskutiert: molekulare Mimikry zwischen EBV-Proteinen wie EBNA-1 und menschlichen Proteinen wie dem Lupus-Autoantigen Ro, Bystander Activation autoreaktiver T-Zellen durch Entzündungssignale aus den infizierten B-Zellen, Epitope Spreading über das anfangs dominante EBV-Antigen hinaus oder polyklonale Antikörperbildung im Zuge der Vermehrung und Aktivierung der befallenen B-Zellen.
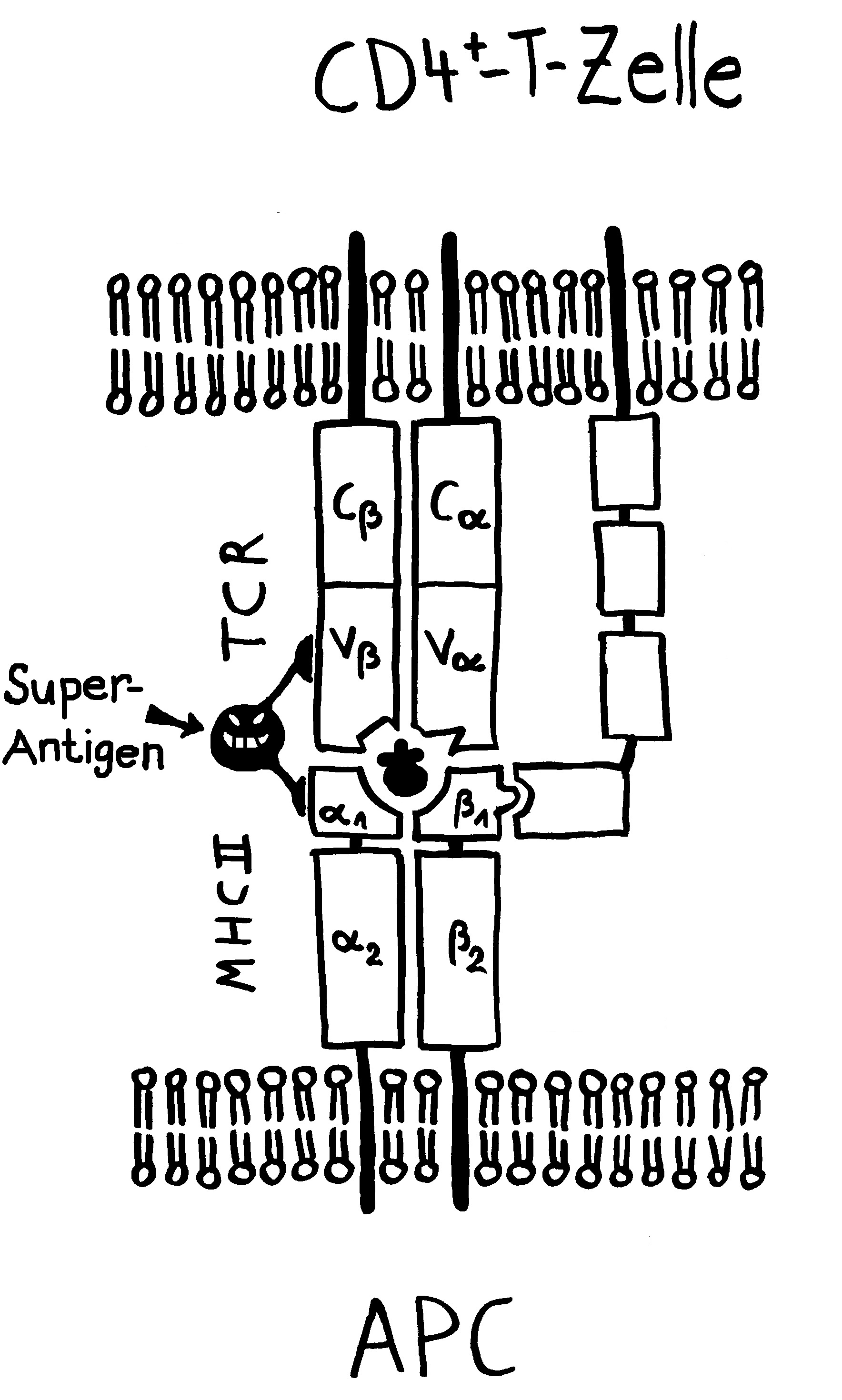
Außerdem wurde spekuliert, die Viren könnten naive autoreaktive B-Zellen so umprogrammieren, dass sie sich auch ohne Aktivierung durch Autoantigen-Kontakt in sehr langlebige Gedächtnis-B-Zellen umwandeln, die dann später Autoimmunreaktionen auslösen. Die Viren könnten auch endogene Retroviren wie HERV-K18 oder HERV-W aktivieren, die normalerweise untätig in unserem Genom schlummern, nach ihrer Erweckung durch EBV aber Superantigene herstellen, die zahlreiche T-Zellen polyklonal aktivieren könnten. Bewiesen ist aber nichts.
Dass im Blut von Patienten mit Autoimmunerkrankungen manchmal deutlich mehr Anti-EBV-Antikörper oder EBV-DNA-Moleküle nachzuweisen sind als bei Gesunden, belegt noch keine Verursachung der Erkrankung durch EBV: Vielleicht stört umgekehrt die Autoimmunerkrankung das Gleichgewicht in den infizierten B-Zellen, sodass die Viren aus ihrem Latenzzustand erwachen und sich vermehren. Da die üblichen Tiermodelle für Autoimmunerkrankungen, insbesondere Mäuse- und Rattenstämme, sich nicht mit EBV infizieren lassen, können auch Tierversuche keine rasche Klärung bringen.