Diesen Kurzvortrag habe ich im Juli beim „Markt der Ideen“ von Extinction Rebellion auf dem Kölner Neumarkt gehalten. Wir, d. h. die Köln-Bonner Regionalgruppe der „Scientists for Future“, hatten dort einen Bücherstand, auf dem wir gute Sachbücher zum Klimawandel vorgestellt haben. Bei einer kleinen Marktrecherche habe ich während der Vorbereitung festgestellt, dass bei Amazon jede Menge wirklich schlechter, irreführender Bücher angeboten werden, zwischen denen die wissenschaftlich korrekten und aktuellen Werke fast untergehen.
Auch hier gilt wieder: Ich halte den Vortrag gerne noch mal; Anfragen bitte an die im Impressum genannte Mail-Adresse. Es gibt auch was zu Lachen dabei, und ich bringe gerne einige gute Bücher zu Reinschnuppern mit.

Die schlechten Werke möchte ich nicht im Einzelnen vorstellen oder zitieren, denn die Kommunikationswissenschaften haben gezeigt, dass dabei fast immer etwas im Gedächtnis haften bleibt – selbst wenn man klar dazu sagt, dass die Aussagen falsch oder fragwürdig sind. Stattdessen schlage ich einfache Regeln vor, um rasch zu erkennen, welche Bücher voraussichtlich nichts taugen.
Diese Faustregeln sind – jede für sich genommen – nicht unfehlbar; zum Beispiel gibt es auch gute Bücher mit reißerischem Titel. In der Summe können sie uns aber vor Fehlkäufen und Zeitverschwendung bewahren. Abgeleitet habe ich sie aus Recherchen zu einigen Dutzend schlechten Büchern im Angebot von Amazon – und aus meinem Hintergrundwissen aus der Buchbranche, in der ich einige Jahrzehnte gearbeitet habe.
 Das erste Warnzeichen: reißerische Buchtitel. Und zugleich eine gute Nachricht: Alle mathematisch möglichen Kombinationen der Schlagworte Lüge, Klima, Verblödung, Öko-Diktatur, Falle, Hysterie und Machenschaft sind inzwischen aufgebraucht. Der Buchmarkt ist gesättigt. ?
Das erste Warnzeichen: reißerische Buchtitel. Und zugleich eine gute Nachricht: Alle mathematisch möglichen Kombinationen der Schlagworte Lüge, Klima, Verblödung, Öko-Diktatur, Falle, Hysterie und Machenschaft sind inzwischen aufgebraucht. Der Buchmarkt ist gesättigt. ?
 Das zweite Warnzeichen: Autoren (übrigens fast immer Männer), die zu allen möglichen Themen Bücher schreiben, also keine Experten in einem bestimmten Sachgebiet sind. Viele von ihnen sind Auftrags- oder Vielschreiber, die ihren Lebensunterhalt mit möglichst schnell produzierten Sachbüchern zu Skandel- und Modethemen verdienen; viele andere sind – gelinde gesagt – Exzentriker oder Esoteriker. So gibt es ein den Klimawandel leugnendes Buch von einem Herrn, der sich früher über die gleichzeitige Existenz von Frühmenschen und Dinosauriern ausgelassen hat.
Das zweite Warnzeichen: Autoren (übrigens fast immer Männer), die zu allen möglichen Themen Bücher schreiben, also keine Experten in einem bestimmten Sachgebiet sind. Viele von ihnen sind Auftrags- oder Vielschreiber, die ihren Lebensunterhalt mit möglichst schnell produzierten Sachbüchern zu Skandel- und Modethemen verdienen; viele andere sind – gelinde gesagt – Exzentriker oder Esoteriker. So gibt es ein den Klimawandel leugnendes Buch von einem Herrn, der sich früher über die gleichzeitige Existenz von Frühmenschen und Dinosauriern ausgelassen hat.
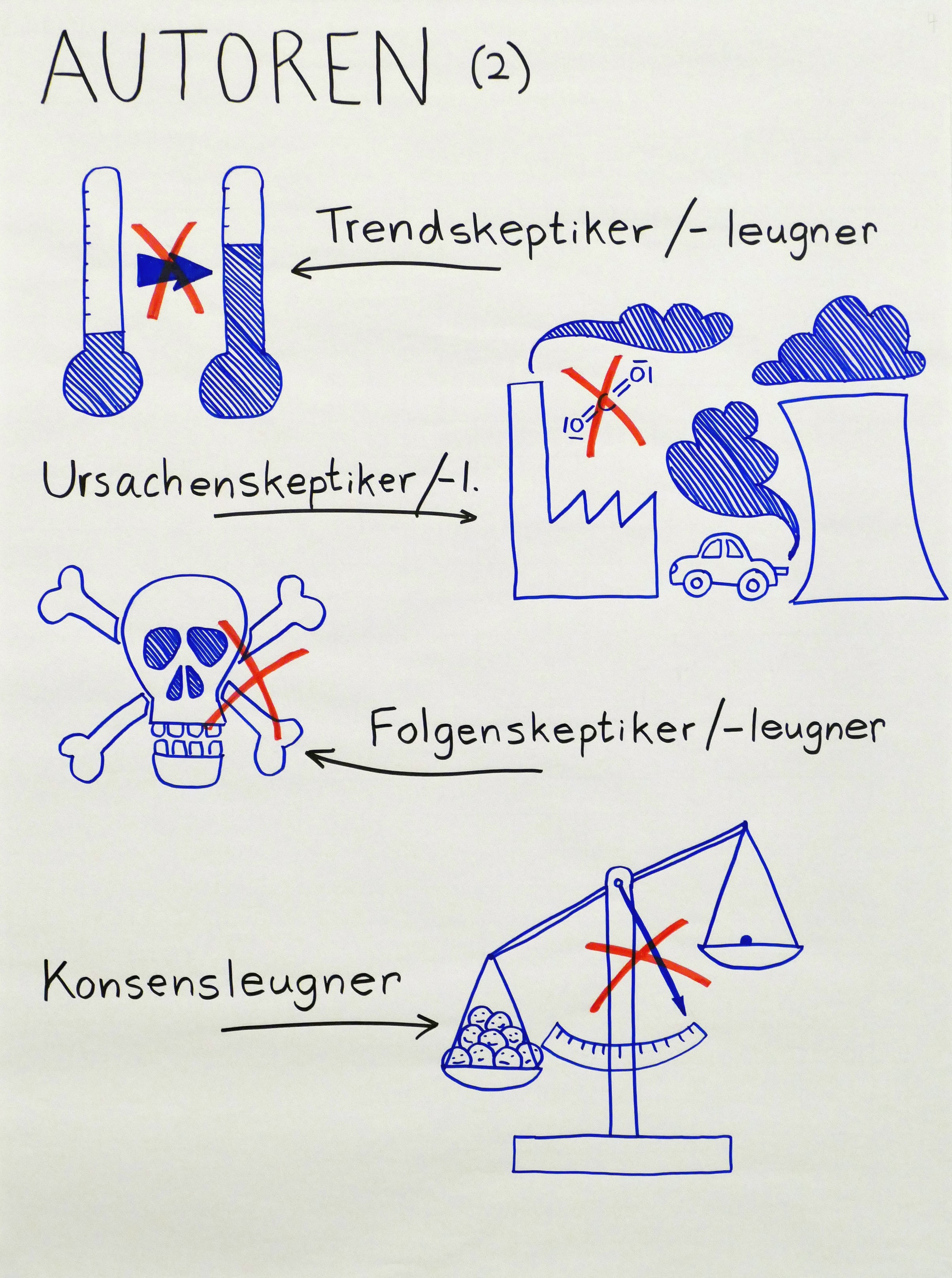 Es sind aber auch Fachleute (im weiteren Sinne, also z. B. Ingenieure) unter den Autoren. Nicht alle leugnen die Tatsache der Erderwärmung an sich: Neben diesen sogenannten Trendskeptikern gibt es auch Ursachenskeptiker (Das Klima wandele sich, aber das liege nicht am Menschen), Folgenskeptiker (Das Klima wandele sich und das liege auch am Menschen, aber es sei nicht schlimm) sowie Konsensleugner (Die Wissenschaft sei sich noch uneins, woran es liege und welche Folgen der Wandel habe).
Es sind aber auch Fachleute (im weiteren Sinne, also z. B. Ingenieure) unter den Autoren. Nicht alle leugnen die Tatsache der Erderwärmung an sich: Neben diesen sogenannten Trendskeptikern gibt es auch Ursachenskeptiker (Das Klima wandele sich, aber das liege nicht am Menschen), Folgenskeptiker (Das Klima wandele sich und das liege auch am Menschen, aber es sei nicht schlimm) sowie Konsensleugner (Die Wissenschaft sei sich noch uneins, woran es liege und welche Folgen der Wandel habe).
Einige Autoren waren früher selbst in der Umweltbewegung aktiv und haben sich nach Auseinandersetzungen abgewandt. Heute arbeiten manche von ihnen für Lobbyorganisationen, die eine Umstellung auf regenerative Energien zu verhindern versuchen.
 Akademische Titel oder Grade sind keine Gewähr für Seriosität. Oftmals haben die Autoren sie in Fachgebieten erworben, die nicht viel mit dem Klima zu tun haben.
Akademische Titel oder Grade sind keine Gewähr für Seriosität. Oftmals haben die Autoren sie in Fachgebieten erworben, die nicht viel mit dem Klima zu tun haben.
 Die Institution, in der oder für die die Autoren arbeiten, kann ein Indiz für die Seriosität sein – wenn man genau hinschaut! „Institut“ ist keine geschützte Bezeichnung; man denke nur an all die „Kosmetik-Institute“. Das Potsdam-Institut für Klimafolgenforschung (PIK) ist eine renommierte Forschungseinrichtung. Das „Institut für Klima und Energie“ (EIKE) ist dagegen eine Briefkasten-Einrichtung: eine Lobby-Institution mit Verbindungen zur AfD. Wenn man sich ein bisschen auskennt, sieht man auch der Website an, dass hier keine seriösen Forscher zu uns sprechen.
Die Institution, in der oder für die die Autoren arbeiten, kann ein Indiz für die Seriosität sein – wenn man genau hinschaut! „Institut“ ist keine geschützte Bezeichnung; man denke nur an all die „Kosmetik-Institute“. Das Potsdam-Institut für Klimafolgenforschung (PIK) ist eine renommierte Forschungseinrichtung. Das „Institut für Klima und Energie“ (EIKE) ist dagegen eine Briefkasten-Einrichtung: eine Lobby-Institution mit Verbindungen zur AfD. Wenn man sich ein bisschen auskennt, sieht man auch der Website an, dass hier keine seriösen Forscher zu uns sprechen.
 Der Verlag, in dem ein Buch erschienen ist, ist kein verlässliches Zeichen für Seriosität: Leider sind auch namhafte Verlage wie Bertelsmann oder Hoffmann und Campe dabei, die sich von sensationalistischen, vermeintlich mutig gegen den Mainstream gerichteten Titeln Umsatz versprechen. Einige der Titel erscheinen in Kleinverlagen aus der rechten bzw. obskurantistischen Ecke, z. B. im FinanzBuch Verlag („Tichys Einblick“).
Der Verlag, in dem ein Buch erschienen ist, ist kein verlässliches Zeichen für Seriosität: Leider sind auch namhafte Verlage wie Bertelsmann oder Hoffmann und Campe dabei, die sich von sensationalistischen, vermeintlich mutig gegen den Mainstream gerichteten Titeln Umsatz versprechen. Einige der Titel erscheinen in Kleinverlagen aus der rechten bzw. obskurantistischen Ecke, z. B. im FinanzBuch Verlag („Tichys Einblick“).
Der Verkaufsrang bei Amazon ist ebenfalls ein problematischen Kriterium. Die Kategorien sind so kleinteilig, dass es nicht schwer ist , mit ein paar gezielten Käufen einen „Bestseller“ zu suggerieren. Gerade wenn ein Buch in den Kreisen von Klimawandel-Leugnern eine gewisse Bekanntheit erlangt hat, kann es auf den vorderen Plätzen landen.
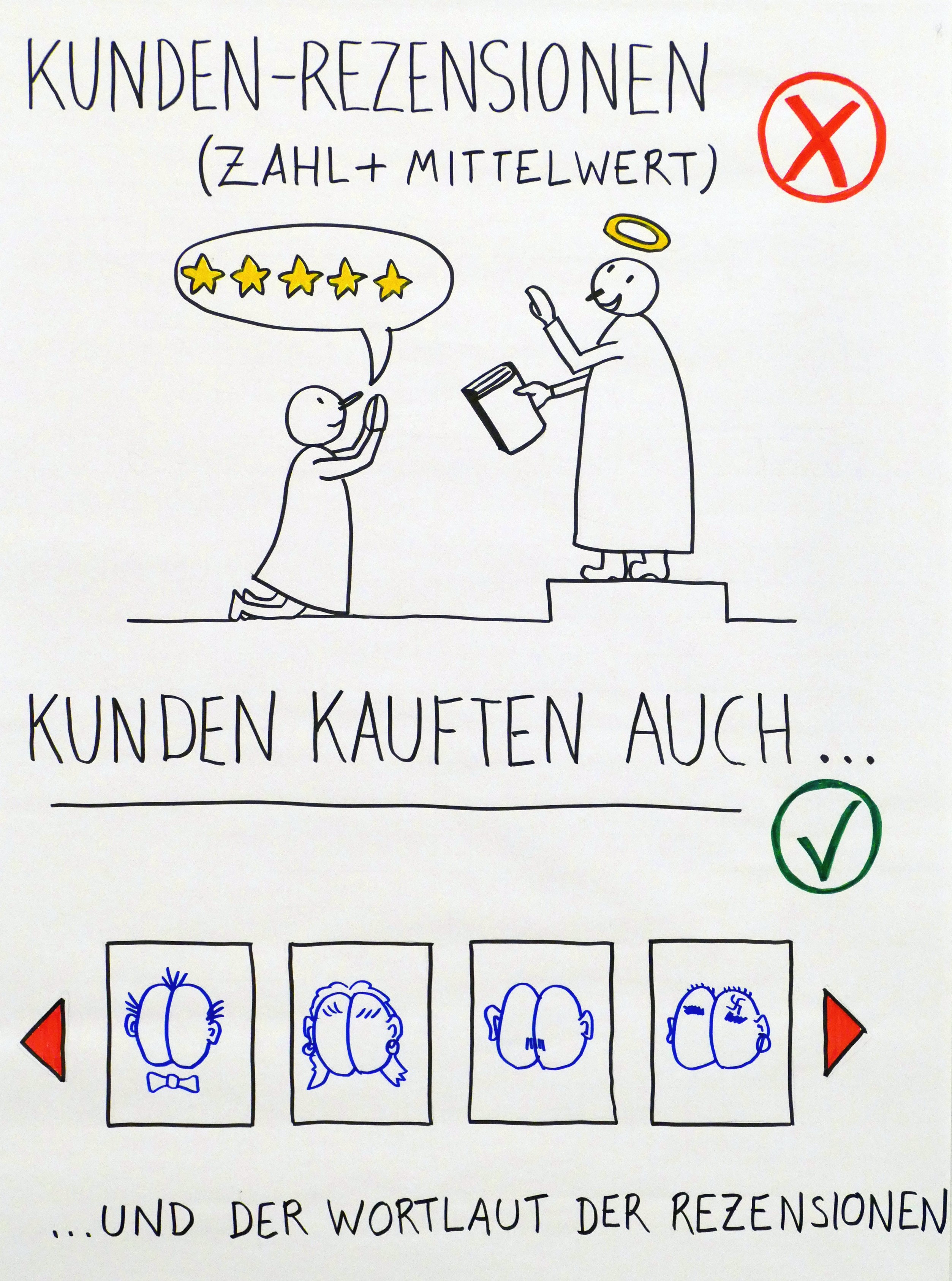 Auch die Zahl und der Mittelwert der Kundenrezensionen sind mit Vorsicht zu genießen: Viele Leser*innen von Büchern, die den Klimawandel leugnen, sind so beseelt von ihrer Mission, dass sie am laufenden Band lobende Besprechungen schreiben – und zugleich Verrisse seröser Werke zum Klimawandel.
Auch die Zahl und der Mittelwert der Kundenrezensionen sind mit Vorsicht zu genießen: Viele Leser*innen von Büchern, die den Klimawandel leugnen, sind so beseelt von ihrer Mission, dass sie am laufenden Band lobende Besprechungen schreiben – und zugleich Verrisse seröser Werke zum Klimawandel.
Ein Blick auf die „Kunden kauften auch“-Rubrik bei Amazon kann dagegen gute Hinweise geben. Wenn da zum Beispiel rechte Autoren auftauchen, spricht das gegen die Seriosität des Klima-Buchs.
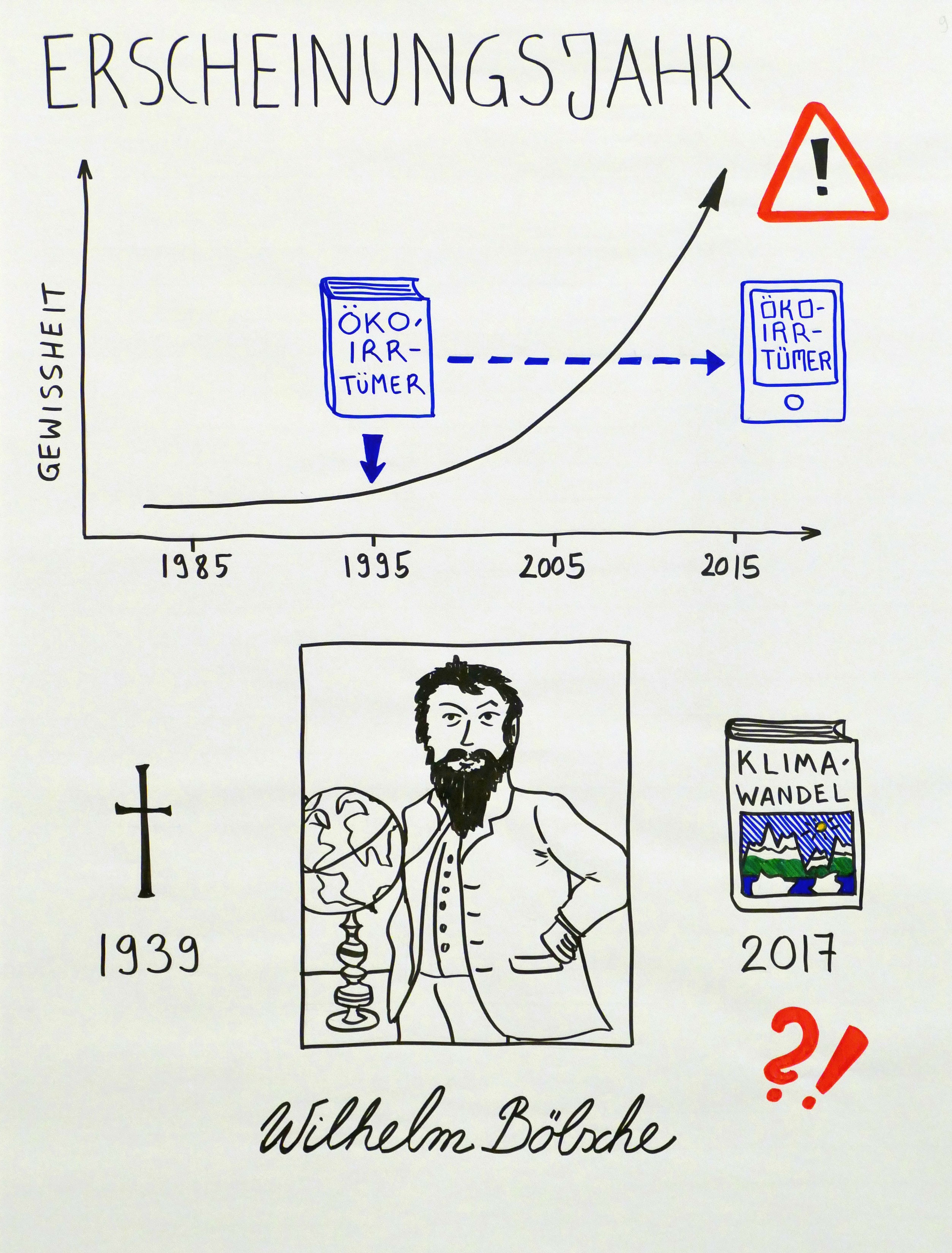
Spielt das Erscheinungsjahr eine Rolle? Ja. Zwar sind die naturwissenschaftlichen Grundlagen des Klimas und des Klimawandels schon länger bekannt. Aber in den letzten Jahren hat es einen enormen Wissenszuwachs gegeben, und auch die politischen, gesellschaftlichen und technologischen Rahmenbedingungen ändern sich rasch. Dass z. B. das „Lexikon der Öko-Irrtümer“ von Miersch und Maxeiner – 1998 erschienen – noch heute unverändert vermarktet wird, ist höchst bedenklich. (Herr Miersch ist inzwischen für die „Deutsche Wildtier Stiftung“ tätig, eine Lobbyorganisation, die den Ausbau regenerativer Energien bekämpft.)
Die alten Schinken werden durchaus noch gelesen, wie neue Kunden-Besprechungen zeigen. Ein Kuriosum zum Abschluss: 2017 erschien ein Text eines bereits 1939 verstorbenen Autors – unter dem Titel „Der erdgeschichtliche Klimawandel: Den wahren Ursachen von Klimaschwankungen auf der Spur“. Der alte Text dient dem Herausgeber als Vehikel für seine eigene Klimawandel-skeptische Botschaft.