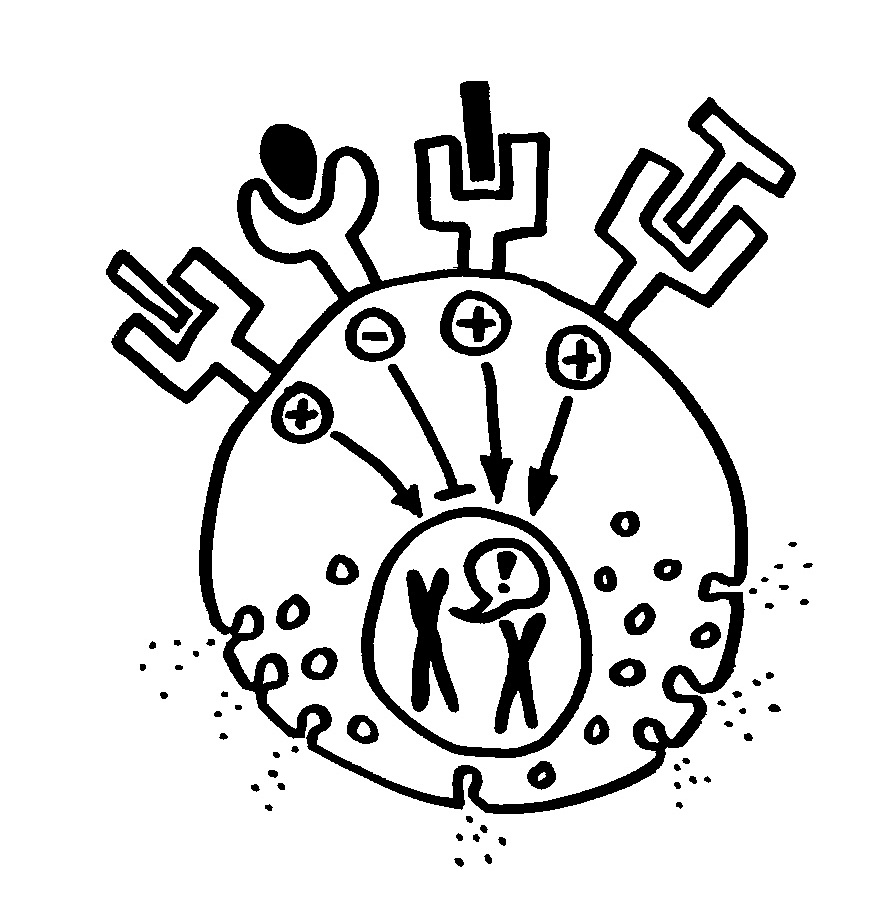
Links: Bei Immunreaktionen vom Typ 1 unterstützen Th1-Zellen die zelluläre Abwehr, mit der vor allem innerzelluläre Pathogene bekämpft werden. Hier beseitigen z. B. Makrophagen bakteriell infizierte Zellen.
Rechts: Bei Immunreaktionen vom Typ 2 helfen Th2-Zellen bei der humoralen Abwehr, die sich vor allem gegen extrazelluläre Pathogene richtet.
Die beiden Arme der Abwehr verstärken sich selbst und hemmen sich gegenseitig (Botenstoff-Luftballons). Aber die Vorstellung, dass immer nur ein Arm aktiv ist, ist überholt: Es gibt kombiniert zellulär-humorale Immunreaktionen.
Sie dürfen diese Zeichnung gerne in Folien etc. übernehmen, sofern Sie die Quelle angeben: Dr. Andrea Kamphuis, https://autoimmunbuch.de