Grundlagenzeichnung, Erläuterung im Buch (und in jedem Bio-Lehrbuch):

Grundlagenzeichnung, Erläuterung im Buch (und in jedem Bio-Lehrbuch):
Mustergültige, klassische Autoimmunerkrankungen erfüllen die Witebsky-Rose-Kriterien:
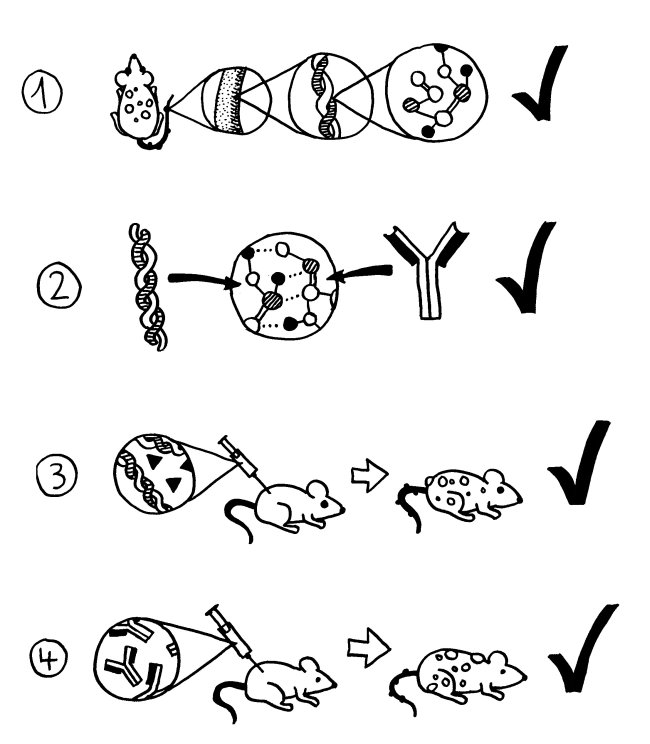
Bei etlichen Störungen, die ziemlich sicher Autoimmunerkrankungen sind, scheitern wir allerdings schon am ersten Kriterium.
Die nächste Zeichnung für Teil 2 des Buchs:
 Antikörper, Komplementfaktoren, Surfactant-Proteine und andere Moleküle des Immunsystems überziehen Krankheitserreger oder Fremdstoffe und machen sie so für Fresszellen wie die Makrophagen attraktiver. Diesen Vorgang nennt man Opsonierung (vom griechischen ópson = Speise).
Antikörper, Komplementfaktoren, Surfactant-Proteine und andere Moleküle des Immunsystems überziehen Krankheitserreger oder Fremdstoffe und machen sie so für Fresszellen wie die Makrophagen attraktiver. Diesen Vorgang nennt man Opsonierung (vom griechischen ópson = Speise).
Zu den ältesten Quellen für das Aufkommen der rheumatoiden Arthritis in Europa zählen Gemälde des Barockmalers Peter Paul Rubens (1577-1640), der vermutlich selbst unter schwerer rheumatoider Arthritis litt – auch wenn er die Krankheit als Gicht bezeichnete.
 In seinem Atelier übernahmen zahlreiche Helfer die Ausführung großer Teile der Gemälde, Aber für die Gesichter und Hände blieb er selbst zuständig, und Kunsthistoriker nehmen an, dass er in seinen Vorstudien oftmals die eigenen Hände porträtierte.
In seinem Atelier übernahmen zahlreiche Helfer die Ausführung großer Teile der Gemälde, Aber für die Gesichter und Hände blieb er selbst zuständig, und Kunsthistoriker nehmen an, dass er in seinen Vorstudien oftmals die eigenen Hände porträtierte.
Das Selbstporträt, auf dem diese Zeichnung basiert, entstand vermutlich um 1638. Zur amerikanischen Herkunft des unbekannten Erregers, der die Autoimmunerkrankung rheumatoide Arthritis mutmaßlich auslöst, habe ich hier früher schon etwas geschrieben.
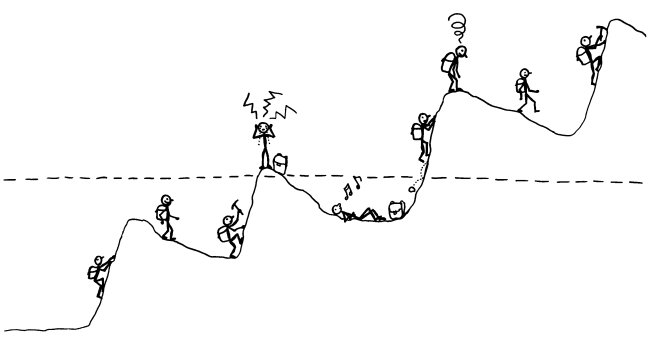 Viele Autoimmunerkrankungen verlaufen schubförmig. Am bekanntesten ist das bei der schubförmig remittierenden Multiplen Sklerose (RR-MS). Aber auch bei Typ-1-Diabetes kann auf den ersten Ausbruch von Symptomen, der zur Diagnose führt, eine Zeit der scheinbaren Genesung folgen – die sogenannte Honeymoon-Phase. Und bei den meisten Autoimmunerkrankungen geht der symptomatischen Phase (oberhalb der gestrichelten Linie) unbemerkt eine langjährige Entgleisung des Immunsystems voran, bei der nach und nach mehr Autoantikörper oder autoreaktive T-Zellen entstehen und es den regulatorischen T-Zellen immer schwerer fällt, diese selbstzerstörerischen Elemente in den Griff zu bekommen.
Viele Autoimmunerkrankungen verlaufen schubförmig. Am bekanntesten ist das bei der schubförmig remittierenden Multiplen Sklerose (RR-MS). Aber auch bei Typ-1-Diabetes kann auf den ersten Ausbruch von Symptomen, der zur Diagnose führt, eine Zeit der scheinbaren Genesung folgen – die sogenannte Honeymoon-Phase. Und bei den meisten Autoimmunerkrankungen geht der symptomatischen Phase (oberhalb der gestrichelten Linie) unbemerkt eine langjährige Entgleisung des Immunsystems voran, bei der nach und nach mehr Autoantikörper oder autoreaktive T-Zellen entstehen und es den regulatorischen T-Zellen immer schwerer fällt, diese selbstzerstörerischen Elemente in den Griff zu bekommen.
Für jeden Aufsatz über das Immunsystem, den ich gelesen habe, wachsen zwei neue nach, die ich unbedingt auch noch lesen muss.

Im Jahr 2011 hatten in den Vereinigten Staaten etwa 11 Millionen Menschen Krebs und nach Schätzung der National Institutes of Health (NIH) etwa 23,5 Millionen Menschen Autoimmunerkrankungen. Die American Autoimmune Related Disease Association (AARDA) geht sogar von etwa 50 Millionen betroffenen Amerikanern aus.
Im Jahr 2003 standen in den USA zur Erforschung von Krebs gut 6 Milliarden US-Dollar zur Verfügung. Die Autoimmunerkrankungen wurden mit knapp 600 Millionen Dollar erforscht. Neuere Zahlen kenne ich nicht.
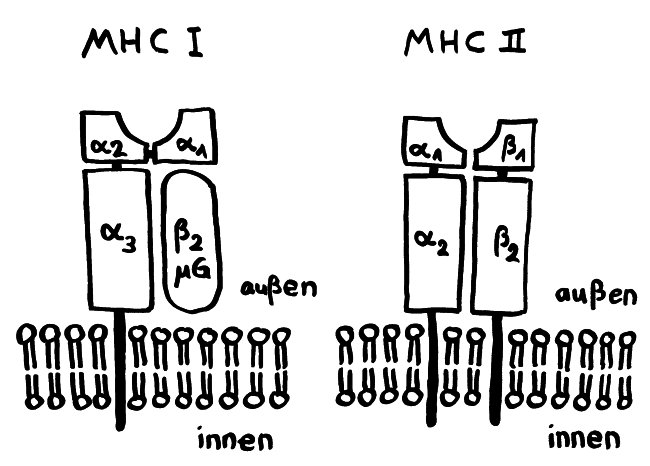
Weitere Grundlagen-Zeichnungen; Erläuterungen folgen im Buch.
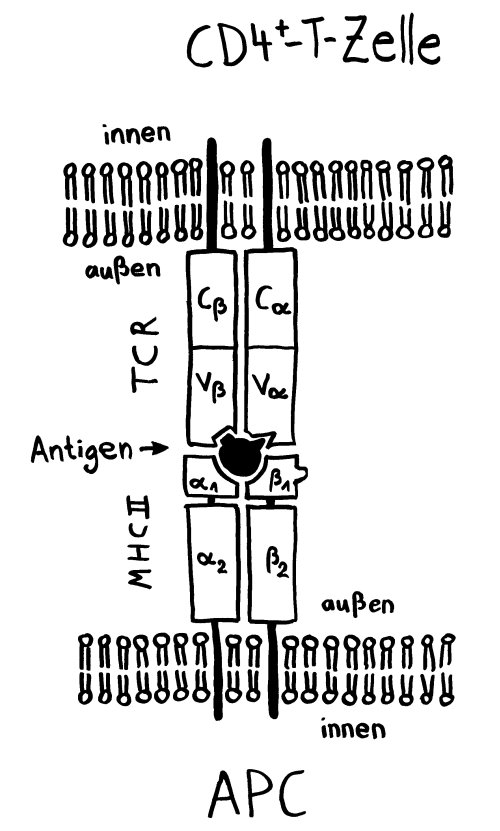
MHC-Klasse-II-Komplex mit Antigen wird von passendem T-Zell-Rezeptor erkannt:
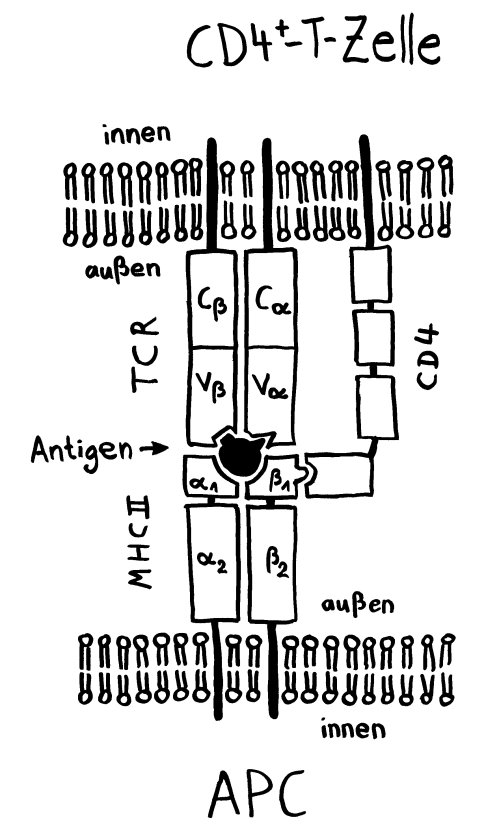
CD4 dockt außen an MHC-II-β-Kette an und stabilisiert die Verbindung:
Superantigene binden sowohl an MHC-Klasse-II-Moleküle als auch an T-Zell-Rezeptoren außerhalb der Antigenbindungsstelle. So lösen sie starke Immunreaktionen in allen möglichen T-Zellen aus:
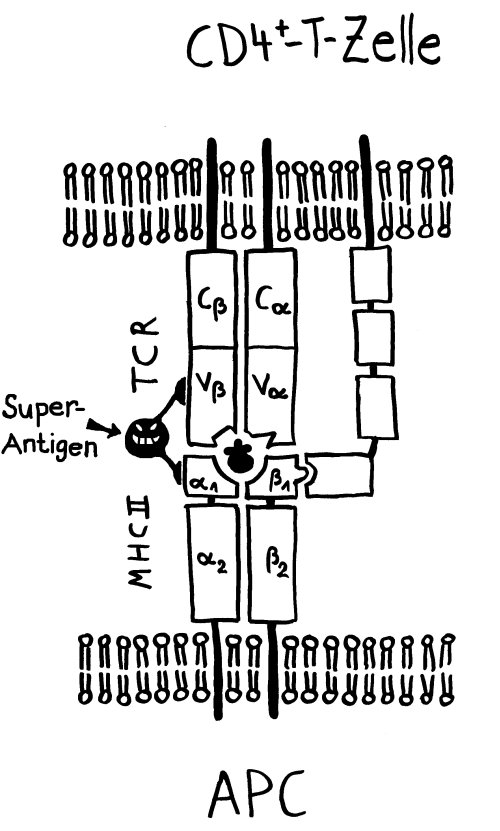
(Quelle für Anordnung Superantigen-Bindung: http://course1.winona.edu/kbates/Immunology/images/figure_11_08.jpg)
Nach und nach arbeite ich die Liste der fehlenden Zeichnungen für die ersten beiden Buchteile ab. Heute sind die Rezeptoren der erworbenen Immunität an der Reihe.
 BCR= B-Zell-Rezeptor, Ig = Immunglobulin, Fc = konstantes Fragment
BCR= B-Zell-Rezeptor, Ig = Immunglobulin, Fc = konstantes Fragment
TCR = T-Zell-Rezeptor, CD3 = Korezeptor des T-Zell-Rezeptors
ITAM = immunoreceptor tyrosine-based activation motif, zuständig für die Signalweiterleitung
Man ist ja dankbar für jeden anschaulichen Wirkstoffnamen.