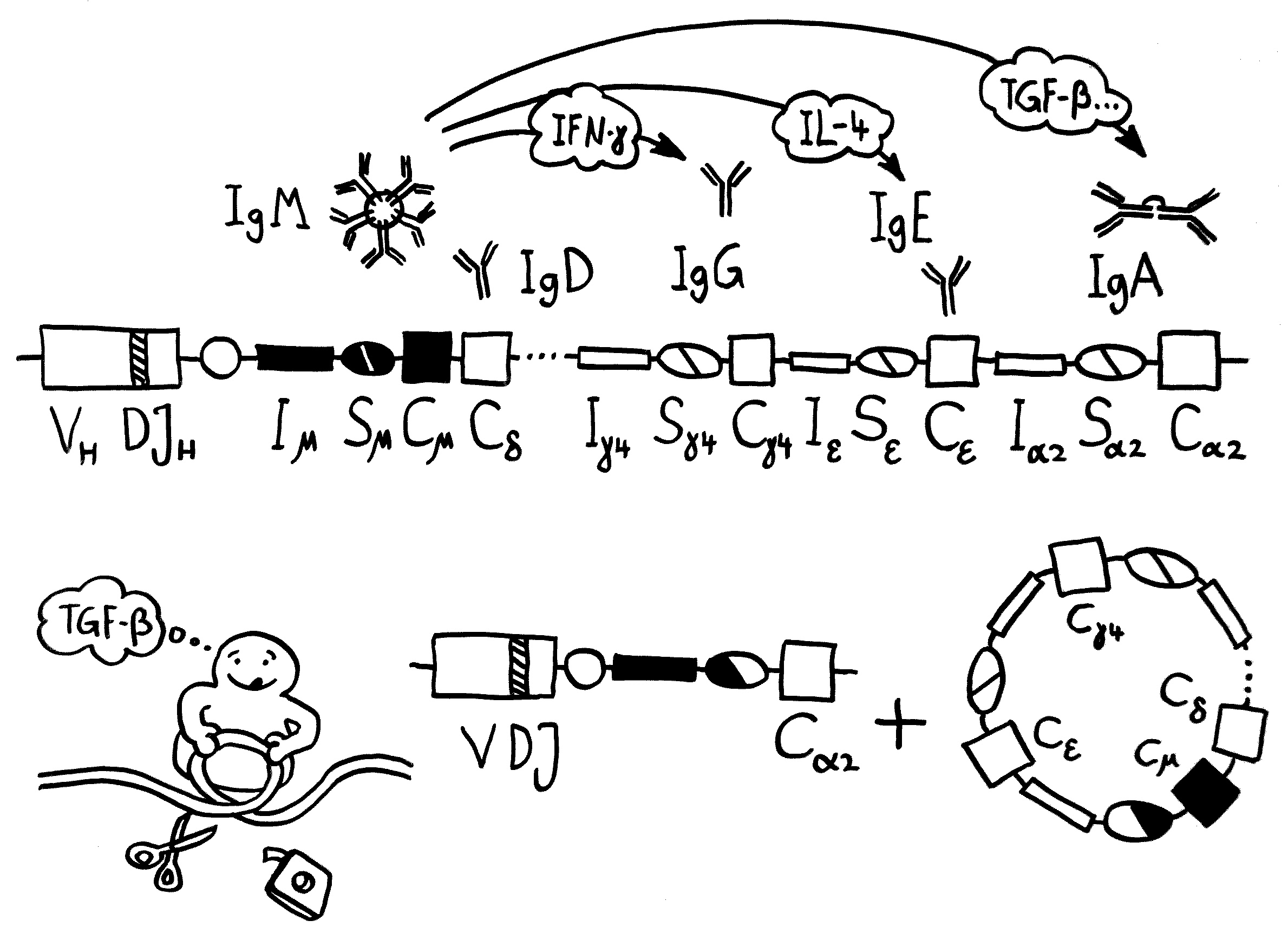
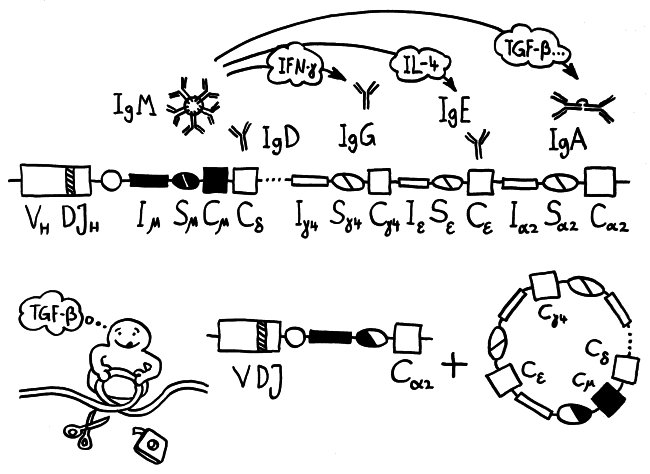
Dass Muttermilch Antikörper enthält, die das Neugeborene in den ersten Monaten vor Infektionen schützen, ist schon länger bekannt. Aber Milch leistet noch mehr für das Immunsystem des Nachwuchses, wie zwei neuere Arbeiten zeigen:
M. K. Ghosh et al. (2016): Maternal Milk T Cells Drive Development of Transgenerational Th1 Immunity in Offspring Thymus (Open Access); dazu auch die Pressemitteilung der Universität: Vaccinating Babies Without Vaccinating Babies
In der Vorläuferstudie hatten die Forscher herausgefunden, dass Mäuse ihrem Nachwuchs beim Säugen nicht nur durch Antikörper, sondern auch durch Immunzellen eine Immunität gegen Pathogene vermitteln, mit denen ihr eigenes Immunsystem kürzlich konfrontiert wurde. Seltsamerweise ist diese Immunität noch beim erwachsenen Nachwuchs nachzuweisen, obwohl dieser keinerlei mütterliche Immunzellen mehr enthält. Die Natur und die Entstehung der Zellen, die diese Immunität vermitteln, sollte hier untersucht werden. Um eine Übertragung im Mutterleib auszuschließen, ließ man die gegen das Bakterium Mycobacterium tuberculosis oder gegen den Pilz Candida albicans immunisierten Mäuseweibchen fremden Nachwuchs aufziehen.
Die Immunität wird offenbar von Gedächtnis-T-Zellen übertragen, die über CD4+-Marker und MHC-Klasse-II-Komplexe verfügen – eine kuriose Kombination, denn normalerweise empfangen CD4+-T-Zellen Signale von antigenpräsentierenden Zellen wie etwa dendritischen Zellen, die Antigene auf MHC-Klasse-II-Komplexen präsentieren. Dendritische Zellen sind aber viel zu kurzlebig, um die hier beobachteten Effekte zu erklären; es waren eindeutig antigenpräsentierende CD4+-T-Zellen, die die Immunität übertrugen – vielleicht, weil nur T-Zellen gezielt in den Thymus wandern können. Wie diese Zellen an die MHC-Klasse-II-Komplexe gelangt sind, ist unklar. Die Autoren vermuten Trogozytose: die Übergabe von Membranflößen einschließlich MHC-Komplex und Kostimulatoren an einer immunologischen Synapse, also einer Bindungsstelle zwischen der (primären) antigenpräsentierenden Zelle und einer T-Zelle, deren T-Zell-Rezeptor spezifisch an den Komplex bindet. Diesen Mechanismus habe ich hier bereits vorgestellt.
Nach der Aufnahme über die Muttermilch wandern diese ungewöhnlichen mütterlichen Gedächtnis-T-Zellen gezielt in den Thymus und die Milz der Mäusebabies. Um an den Grenzen – also am Brustdrüsen-, Darm- und Thymusepithel – nicht von anderen Immunzellen aufgehalten zu werden, „verschlucken“ sie vermutlich ihre MHC-Klasse-II-Komplexe samt Antigenen in Vesikeln und befördern sie erst am Ziel wieder an die Zelloberfläche. Im Thymus werden die MHC-Klasse-II-Komplexe einschließlich der Antigene womöglich durch eine weitere Trogozytose an „ordentliche“ antigenpräsentierende Zellen übergeben, oder die CD4+-T-Zellen werfen die Antigene ab, und antigenpräsentierende Zellen nehmen sie auf.
Jedenfalls werden die Antigene aus den Pathogenen, mit denen die Mütter infiziert waren, nun den unreifen Mäusebaby-Thymozyten präsentiert, die daraufhin zu CD8+-T-Zellen mit einer Spezifität für diese Antigene heranreifen. Diese Immunitätsübertragung nennen die Autoren „maternal educational immunity“, um sie von der passiven Immunität zu unterscheiden, die vor allem durch mütterliche Antikörper in der Milch übertragen wird und sich rasch verliert, da diese Antikörper im Jungtier nicht nachproduziert werden können.
In der Pressemitteilung der Universität finden sich interessante Spekulationen über eine mögliche Nutzung dieses Mechanismus zur „indirekten Impfung“ von Säuglingen (nämlich durch Impfung der Mütter während der Schwangerschaft) und über die hohe historische Überlebensrate von Kleinkindern aus Adelsfamilien, die häufig von Ammen aus der Unterschicht gestillt wurden und so vielleicht eine besonders gute „Immunsystem-Erziehung“ genossen. Dabei sollte aber nicht vergessen werden, dass die Reifung des Immunsystems bei jungen Mäusen anders verläuft als bei Menschenkindern.
M. A. Koch et al. (2016): Maternal IgG and IgA Antibodies Dampen Mucosal T Helper Cell Responses in Early Life (Bezahlschranke, nur Abstract und eine Abbildung); dazu auch Meldung „Breast Milk Primes Gut for Microbes“ in The Scientist
Mütterliche, über die Milch übertragene Antikörper der Typen IgG und IgA dienen vor allem dazu, Pathogene im Darm junger Mäuse zu bekämpfen, solange deren Immunsystem dazu noch nicht imstande ist – so glaubte man bisher. Jetzt zeigt sich, dass insbesondere IgG auch Immunreaktionen hemmt, und zwar solche gegen nützliche Bakterien, die nach der Geburt den Darm von Mäusebabies besiedeln. Fehlen die mütterlichen Antikörper, reagiert das Lymphgewebe am Darm heftig auf die neue Darmflora: Es entstehen viel mehr T-Helferzellen, die wiederum B-Zellen zur Produktion von Antikörpern gegen die gutartigen Darmbakterien anregen.
Allerdings scheinen die Mäuse, denen das mütterliche IgG vorenthalten wurde, keine langfristigen Gesundheitsschäden davonzutragen. Der Begleitartikel in The Scientist stellt dennoch Spekulationen über langfristige Folgen einer gestörten Mikrobiom-Entwicklung an, etwa Morbus Crohn und Colitis ulcerosa – nur um dann abzuwiegeln und auf die Unterschiede zwischen Mensch und Maus hinzuweisen. Zum Beispiel darauf, dass menschliche Muttermilch viel weniger IgG enthält als die von Mäusen. Es ist zum Mäusemelken.