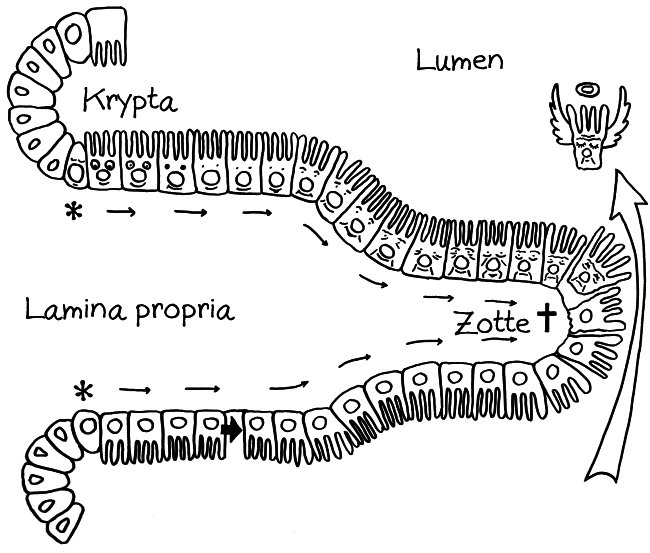
In der Fachliteratur zum Mikrobiom ist immer wieder von Resilienz die Rede, also der Rückstellkraft eines dynamischen Systems bei Störungen. Wie in der Makroökologie, die sich mit Lebensräumen wie Wäldern beschäftigt, zeigt sich auch bei der Erforschung des Mikrobioms: Ein Ökosystem kehrt nach einer Störung umso sicherer in seinen ursprünglichen Gleichgewichtszustand zurück, je größer seine Biodiversität ist – je mehr Arten also in ihm vorkommen.
 Das Rückstellvermögen eines Systems wird gerne durch eine Kugel in einem parabelförmigen Tal versinnbildlicht, das bei hoher Resilienz tief und bei geringer Resilienz flach ist. Eine Störung, etwa eine akute Erkrankung, führt in einem bereits „erodierten“, zum Beispiel durch mehrfachen Antibiotika-Einsatz oder einseitige Ernährung verarmten System leichter dazu, dass die Kugel aus ihrer Kuhle herauskullert.
Das Rückstellvermögen eines Systems wird gerne durch eine Kugel in einem parabelförmigen Tal versinnbildlicht, das bei hoher Resilienz tief und bei geringer Resilienz flach ist. Eine Störung, etwa eine akute Erkrankung, führt in einem bereits „erodierten“, zum Beispiel durch mehrfachen Antibiotika-Einsatz oder einseitige Ernährung verarmten System leichter dazu, dass die Kugel aus ihrer Kuhle herauskullert.
Die größere Widerstandskraft artenreicher Ökosysteme ist vermutlich auf ihre sogenannte funktionelle Redundanz zurückzuführen: Jede für den Erhalt des Ganzen wichtige Aufgabe – beispielsweise die Herstellung eines Stoffwechselprodukts, das andere Arten im Mikrobiom oder die Zellen des Wirts benötigen – kann durch mehrere Angehörige des Systems erledigt werden.

Wie bei einem Klappbuch, in dem Kinder aus mehreren Köpfe, Rümpfen und Fortbewegungsorganen alle möglichen Fabelwesen zusammensetzen können, macht es nicht viel aus, wenn ein Element verloren geht: Reißt man ein Kopf-Blatt heraus, sind immer noch andere Köpfe übrig, sodass das Wesen komplett bleibt. Gibt es dagegen nur ein Kopf-Blatt, steht und fällt die Fähigkeit, ganze Wesen zu bilden, mit dem Erhalt dieses einen Blattes.
Angenommen, unser System könne zwei Gleichgewichtszustände annehmen, einen „gesunden“ (links) und einen „kranken“ (rechts), und befinde sich am Anfang in einem gesunden dynamischen Gleichgewicht (Homöostase, Kugel links). Wie kommt es dann zu chronischen Erkrankungen?
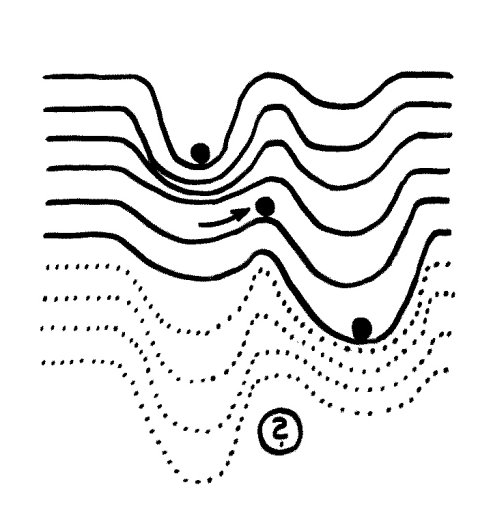
Wenn z. B. unsere Darmflora durch die Ernährung oder den ständigen Einsatz antimikrobieller Substanzen allmählich verarmt, schwindet ihre Resilienz. Das linke Tal wird flacher, sodass eine Störung wie eine Infektion die Kugel ins rechte Tal schubsen kann (Dysbiose). Um die Homöostase wiederherzustellen, reicht es meist nicht, die Kugel noch einmal anzustupsen, denn der Abhang, den sie überwinden müsste, um wieder nach links zu gelangen, ist zu steil.
Von langfristig eingenommenen pro- oder präbiotischen Lebensmitteln oder radikaleren Mikrobiom-Therapien wie der Verabreichung lebender Würmer oder Stuhltransplantationen erhofft man sich stattdessen einen allmählichen Umbau des „Landschaftsprofils“: Das linke Tal soll wieder tiefer und das rechte flacher werden, sodass das System in seinen gesunden Gleichgewichtszustand zurückspringen kann und dann auch dort bleibt.
Dass das gelingt, ist keineswegs sicher: Die Diversität und damit die Resilienz eines Ökosystems ist viel leichter zu reduzieren als wieder aufzubauen. Das gilt für Urwälder und Fichten-Monokulturen ebenso wie für unsere Darmflora.